Таблица 3.3
Сравнительный анализ плотности рыбного населения на участках мелководной поймы с затопленным лесом (над чертой) и без него (под чертой).
|
m |
n |
s |
S |
С |
P |
|
|
день |
2.1 |
19 |
2.1 |
1.0 |
101.7% |
<0.01 |
|
0.3 |
305 |
1.2 |
0.1 |
356.7% |
||
|
ночь |
6.5 |
27 |
4.0 |
1.5 |
61.7% |
<0.01 |
|
2.8 |
312 |
4.5 |
0.5 |
163.7% |
Примечание. Обозначения и единицы измерения как в Таблице 1, в последнем столбце достоверность различий в плотности (t-тест).
Данные результаты позволяют сделать заключение, что трехмерные субстраты в пресных водоемах, имеющие сложную внутреннюю структуру подобную морским рифам, могут выполнять и сходную с ними функцию (Герасимов, Комова, 1987; Герасмов, Слынько, 1990; Герасимов, Поддубный, 1999). В данном случае они способствуют увеличению емкости среды и устойчивости населяющих их сообществ гидробионтов.
Вышеуказанные факторы стабильно определяют перераспределение рыб между отдельными биотопами Рыбинского водохранилища на протяжении всего времени его существования.
Общую картину распределения рыб на акватории водохранилища на современном этапе в основном определяют два фактора: уровень изъятия рыбы из водохранилища (легальный и нелегальный) и динамика численности нового для водохранилища вида – каспийской тюльки (Экологические проблемы Верхней Волги, 2001).
Первый из указанных факторов определяет картину распределения демерсальных рыб. В настоящий период в результате перелова наблюдается значительное снижение уловов (коэффициент регрессии r = -0.54 за период с 1970 по 1990 гг., r = -0.94 – с 1990 по 2002 гг.). Происходит сокращение численности крупных особей в популяциях основных промысловых рыб Рыбинского водохранилища. Это видно и по результатам контрольных обловов и по данным эхосъемок (рис.3.2, коэффициент регрессии = - 0.75; F=29.1; p=0.01). В 2002 году средняя ихтиомасса (по данным гидроакустических съемок) за все время существования водохранилища опустилась ниже отметки 100 кг/га (рис. 3.3). Тогда как за период с 1978 до 1999 года средняя ихтиомасса составляла 142 ± 11 кг/га (коэффициент регрессии = - 0.25; F=0.82; p=0.38; доверительный интервал 131-153 кг/га при 95% вероятности).
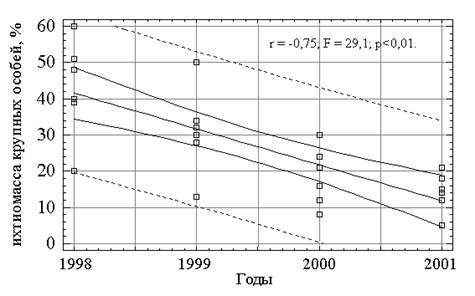
Рис. 3.2. Динамика ихтиомассы крупных особей (по данным эхосъемок, в % к общей ихтиомассе).
В результате подобного воздействия изменилась общая картина распределения рыб. Отмечено изменение пространственной структуры: уменьшилось количество скоплений рыб с высокой плотностью, уменьшились размеры скоплений и расстояния между скоплениями. На это указывают и показатели агрегированности (рис.3.3). До начала 90-х среднее значение коэффициента агрегированности Ллойда составляло 10.12 ± 2.31, а после 2000 года оно уменьшилось в три раза и составило 3.68 ± 1.34.
![]()
Рис. 3.3. Средняя ихтиомасса (кг/га) за период с 1978 по 2003 годы по данным гидроакустических съемок. Цифры над гистограммами – значение коэффициента агрегированности рыб по Ллойду (Lloyd, 1967).
На то, что снижение пространственной сегрегации рыб может быть связано с усилением пресса рыболовства указывается и другими исследователями (O’Driscoll et al., 2000; Blanchard, 2001).
Распределение рыб пелагического комплекса не связано с выловом, поскольку ни один из основных его видов не используется промыслом. Численность и встречаемость пелагических видов связана с динамикой численности вида - доминанта – каспийской тюльки.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.