При этом, если ток течет от электрода,
находящегося у внешней поверхности мембраны, к электроду, находящемуся в
цитоплазме у внутренней поверхности (I>0),
потенциал мембраны будет уменьшаться по абсолютной величине. В этом случае
говорят о деполяризации мембраны. Если же направление тока противоположно (I<0), то абсолютное значение мембранного потенциала
будет увеличиваться. Эта 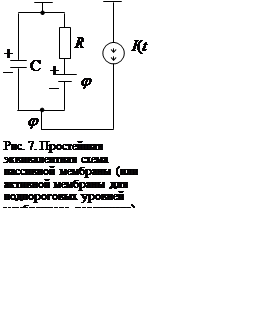 ситуация характеризуется термином
«гиперполяризация». Выражение для мембранного потенциала (16) справедливо лишь
для интервалов времени, не превышающих длительность импульса воздействующего
тока, а также постоянстве сопротивления мембраны, т.е. в отсутствие нелинейных
(R¹R(j))
или параметрических (R¹R(t)) эффектов. Эти условия в большей степени выполняются
для гиперполяризирующего импульса. Мембрана ведет себя при этом подобно
пассивной RC цепи с постоянной времени t = RC.
Форма импульса потенциала на ней будет изменяться от линейно спадающей (при
ситуация характеризуется термином
«гиперполяризация». Выражение для мембранного потенциала (16) справедливо лишь
для интервалов времени, не превышающих длительность импульса воздействующего
тока, а также постоянстве сопротивления мембраны, т.е. в отсутствие нелинейных
(R¹R(j))
или параметрических (R¹R(t)) эффектов. Эти условия в большей степени выполняются
для гиперполяризирующего импульса. Мембрана ведет себя при этом подобно
пассивной RC цепи с постоянной времени t = RC.
Форма импульса потенциала на ней будет изменяться от линейно спадающей (при ![]() ) до прямоугольной (при
) до прямоугольной (при ![]() ). При отключении возбуждающего тока
(
). При отключении возбуждающего тока
(![]() ) потенциал будет возвращаться к исходному
состоянию (
) потенциал будет возвращаться к исходному
состоянию (![]() ) по экспоненциальному закону (рис.
6). Такая же картина будет наблюдаться на мембранах невозбудимых клеток и в
случае деполяризующего тока, а также на мембранах возбудимых, если уровень
потенциала не достигает порогового значения.
) по экспоненциальному закону (рис.
6). Такая же картина будет наблюдаться на мембранах невозбудимых клеток и в
случае деполяризующего тока, а также на мембранах возбудимых, если уровень
потенциала не достигает порогового значения.
Характер изменения потенциала мембраны
возбудимых клеток существенно изменяется, если под воздействием внешнего
источника тока он достигнет порогового значения. В этом случае потенциал перестает
зависеть от воздействующего тока: происходит его быстрый рост до максимального
значения ![]() и спад к уровню потенциала покоя, в
некоторых случаях носящий колебательный характер с большим декрементом
затухания (потенциал может опуститься ниже уровня
и спад к уровню потенциала покоя, в
некоторых случаях носящий колебательный характер с большим декрементом
затухания (потенциал может опуститься ниже уровня ![]() ,
гиперполяризоваться и далее плавно к нему вернуться (рис. 6)). Такого рода
поведение функции потенциала связывается с резким потенциалзависимым изменением
проводимости одного из типов ионных каналов (в случае нервных и мышечных клеток
– натриевых).
,
гиперполяризоваться и далее плавно к нему вернуться (рис. 6)). Такого рода
поведение функции потенциала связывается с резким потенциалзависимым изменением
проводимости одного из типов ионных каналов (в случае нервных и мышечных клеток
– натриевых).
Причем увеличение проводимости
мембраны пропорционально смещению ее потенциала относительно потенциала покоя,
вследствие чего в системе возникает положительная обратная связь: увеличение
проводимости приводит к увеличению смещения потенциала, а смещение потенциала –
к увеличению проводимости. Это обусловливает быстрый регенеративный процесс,
приводящий к установлению на мембране потенциала, близкого к стационарному потенциалу
данного иона. В самом деле, в соответ соответствии с (8), если проводимость
какого-либо типа ионов (например, k-того) превышает
проводимость всех остальных, то, пренебрегая в знаменателе (8) всеми
проводимостями за исключением ![]() , можно записать
выражение для
, можно записать
выражение для ![]() в виде
в виде 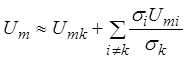 . Из последнего при
. Из последнего при ![]() и следует
и следует ![]() .
.
Проводимость мембраны в состоянии возбуждения зависит также от времени. Зависимость эта носит экстремальный характер: в начале возбуждения проводимость растет, достигает максимума, а в дальнейшем уменьшается до стационарного значения.
Основные результаты теории
возбуждения клеточных мембран были достигнуты при исследовании гигантского
аксона кальмара. Поэтому рассмотрим механизмы генерации потенциала действия
именно на этом примере. Как уже упоминалось, в состоянии покоя потенциал
мембраны нервного аксона составляет величину около –70 мВ. Проницаемости ионных
каналов калия, натрия и хлора при этом соотносятся как 1:0,04:0,45. Как видно,
в состоянии покоя наибольшей проницаемостью обладают калиевые каналы,
вследствие чего потенциал мембраны в соответствии с (8) в основном определяется
калиевой проводимостью. При воздействии на мембрану внешним импульсом тока
деполяризующего направления происходит уменьшение абсолютного значения
потенциала на ней. Это приводит к изменению суммарной (интегральной) проницаемости
мембранных каналов. Наибольшим изменениям оказываются подвержены натриевые
каналы. Ионы натрия по градиенту концентрации поступают внутрь клетки, еще
более понижая потенциал мембраны. В свою очередь, понижение потенциала приводит
к дельнейшему увеличению проницаемости мембраны для ионов натрия. Таким
образом, реализуется сильная положительная обратная связь, приводящая к
возникновению регенеративного процесса. По мере насыщения проводимости
натриевых каналов, а также вследствие увеличения вероятности открывания
калиевых, переносящих положительный заряд из цитоплазмы в окружающую среду,
происходит уменьшение скорости роста мембранного потенциала. Вследствие
взаимодействия двух конкурирующих процессов переноса (Na+
- внутрь, К+ - наружу) потенциал мембраны достигает максимального
значения, близкого к мембранному потенциалу натрия ![]() ,
и составляет величину около 40 мВ. Проницаемости мембраны в этом случае для
калия, натрия и хлора соотносятся как 1:20:0,45. Таким образом, на пике возбуждения
проницаемость натриевых каналов увеличивается в 500 раз в сравнении с
состоянием покоя. Это и проводит к тому, что основной вклад во взвешенный
суммарный потенциал мембраны вносит натриевая компонента.
,
и составляет величину около 40 мВ. Проницаемости мембраны в этом случае для
калия, натрия и хлора соотносятся как 1:20:0,45. Таким образом, на пике возбуждения
проницаемость натриевых каналов увеличивается в 500 раз в сравнении с
состоянием покоя. Это и проводит к тому, что основной вклад во взвешенный
суммарный потенциал мембраны вносит натриевая компонента.
После достижения максимального
значения спад потенциала мембраны связан с уменьшением интегральной
проницаемости натриевых каналов (проницаемость калиевых сохраняет высокий уровень
в течение более длительного времени). Действие положительной обратной связи
опять приводит к регенеративному механизму изменения потенциала мембраны.
Процесс возвращения к исходному уровню, называющийся реполяризацией, носит
колебательный характер, хотя и имеет большой декремент затухания, так что
отклонение от установившегося значения (![]() )
носит характер единичной полуволны гиперполяризации (рис. 6). Она обусловлена
тем, что в этот интервал времени калиевая проводимость еще превышает уровень,
соответствующий состоянию покоя, постоянно релаксируя к нему.
)
носит характер единичной полуволны гиперполяризации (рис. 6). Она обусловлена
тем, что в этот интервал времени калиевая проводимость еще превышает уровень,
соответствующий состоянию покоя, постоянно релаксируя к нему.
Восстановление исходных разностей концентраций, нарушенных (хотя и слабо) в процессе возбуждения, осуществляется с помощью активного (против градиентов) транспорта ионов, осуществляемого ионными насосами.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.