 Пик R
соответствует первой стадии возбуждения – активации натриевых каналов. Пик S обусловлен спадом напряжения на мембране в стадии начальной
быстрой реполяризации. В интервале относительного постоянства мембранного
потенциала ток в межклеточной жидкости равен нулю. В четвертой стадии
производная потенциала отрицательна, что отражается пиком Т.
Пик R
соответствует первой стадии возбуждения – активации натриевых каналов. Пик S обусловлен спадом напряжения на мембране в стадии начальной
быстрой реполяризации. В интервале относительного постоянства мембранного
потенциала ток в межклеточной жидкости равен нулю. В четвертой стадии
производная потенциала отрицательна, что отражается пиком Т.
4.2. Особенности функционирования сердца
 Основной функцией сердца является
насосная, т.е. обеспечение непрерывной перекачки крови по замкнутой системе
кровообращения. Реализуется эта функция за счет способности сердечной мышцы сокращаться
в ответ на электрическое возбуждение. Особенностью сердца млекопитающих
(человека, в частности) является то, что оно может работать, будучи изолированным
от остальной части тела, т.е. автономно. Этот феномен объясняется
существованием в сердце специальных мышечных клеток, именуемых пейсмейкерами (pasemaker). Эти клетки непрерывно генерируют импульсы,
распространение которых приводит к возбуждению сократительных мышц сердца.
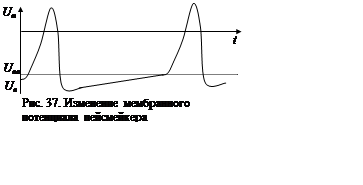
После формирования очередного импульса возбуждения потенциал мембраны
пейсмейкера не устанавливается на уровне потенциала покоя, а медленно
повышается вследствие сохранения механизма обратной связи между изменениями
потенциала мембраны и проводимостью натриевых каналов. Постоянная времени цепи
обратной связи в этом интервале велика, что и объясняет медленное развитие процесса
повышения потенциала. Тем не менее, в некоторый момент он все же достигает
порогового уровня, вслед за чем происходят быстрый регенеративный процесс
деполяризации мембраны и генерация потенциала действия (рис. 37).
Основной функцией сердца является
насосная, т.е. обеспечение непрерывной перекачки крови по замкнутой системе
кровообращения. Реализуется эта функция за счет способности сердечной мышцы сокращаться
в ответ на электрическое возбуждение. Особенностью сердца млекопитающих
(человека, в частности) является то, что оно может работать, будучи изолированным
от остальной части тела, т.е. автономно. Этот феномен объясняется
существованием в сердце специальных мышечных клеток, именуемых пейсмейкерами (pasemaker). Эти клетки непрерывно генерируют импульсы,
распространение которых приводит к возбуждению сократительных мышц сердца.
После формирования очередного импульса возбуждения потенциал мембраны
пейсмейкера не устанавливается на уровне потенциала покоя, а медленно
повышается вследствие сохранения механизма обратной связи между изменениями
потенциала мембраны и проводимостью натриевых каналов. Постоянная времени цепи
обратной связи в этом интервале велика, что и объясняет медленное развитие процесса
повышения потенциала. Тем не менее, в некоторый момент он все же достигает
порогового уровня, вслед за чем происходят быстрый регенеративный процесс
деполяризации мембраны и генерация потенциала действия (рис. 37).
Период следования импульсов возбуждения, вырабатываемых отдельными пейсмейкерами, однако, весьма нестабилен: он может изменяться в процессе работы в пределах двух-трех раз даже в условиях поддержания параметров окружающей среды в высокой степени стабильности. Это связано с тем обстоятельством, что процесс спонтанного повышения потенциала мембраны пейсмейкера в паузу между двумя очередными актами возбуждения происходит весьма медленно, т.е. разность между пороговым потенциалом и потенциалом мембраны на значительном временном интервале изменяется несущественно. Вследствие этого случайное открывание или закрывание какой-то части каналов мембраны может привести к более раннему или позднему началу регенеративного процесса возбуждения мембраны. Наблюдаемая же на практике довольно высокая стабильность частоты сердечных сокращений связана с их коллективным действием из-за наличия связей между отдельными клетками – пейсмейкерами. Более быстрая деполяризация какой-либо части клеток или отдельной клетки приводит к появлению тока ионов между этими клетками и клетками, потенциал которых имеет более низкий уровень. Это замедляет процесс деполяризации одних клеток и ускоряет этот процесс в других. В результате отклонения частоты колебаний от некоторого среднего значения уменьшаются.
Оценим приближенно зависимость нестабильности частоты от числа клеток – возбудителей ритма. По аналогии со случаем токов ионных каналов мембраны (см. с. 9) суммарный ток совокупности клеток-пейсмейкеров равен сумме токов отдельных клеток
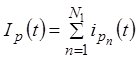 .
.
Здесь ![]() - суммарный ток всех пейсмейкеров в
определенный момент времени,
- суммарный ток всех пейсмейкеров в
определенный момент времени, ![]() - ток отдельного
пейсмейкера в этой же временной точке, N1
– общее число пейсмейкеров. Повторив рассуждения, приведенные при исследовании
тока клеточной мембраны, получим:
- ток отдельного
пейсмейкера в этой же временной точке, N1
– общее число пейсмейкеров. Повторив рассуждения, приведенные при исследовании
тока клеточной мембраны, получим:
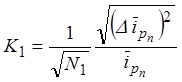 ,
,
где K1 –
относительная флуктуация тока коллектива пейсмейкеров, ![]() -
отклонение тока отдельных пейсмейкеров от среднего значения, соответствующего
данному моменту времени,
-
отклонение тока отдельных пейсмейкеров от среднего значения, соответствующего
данному моменту времени, ![]() - средний ток
конкретного пейсмейкера, соответствующий этому же моменту времени. Если
предположить, что из-за связи между клетками частоты колебаний их мембранного
потенциала не слишком сильно отличаются и максимальное отклонение тока в каждый
момент времени приблизительно равно среднему току в этот момент, т.е.
- средний ток
конкретного пейсмейкера, соответствующий этому же моменту времени. Если
предположить, что из-за связи между клетками частоты колебаний их мембранного
потенциала не слишком сильно отличаются и максимальное отклонение тока в каждый
момент времени приблизительно равно среднему току в этот момент, т.е. ![]() , то формула для K1
получает тот же вид, что и в случае для тока клеточной мембраны:
, то формула для K1
получает тот же вид, что и в случае для тока клеточной мембраны:
![]() .
.
Параметр K1 определяет не только отклонение тока в какой-либо момент интервала формирования мембранного потенциала (например, в максимуме) от среднего значения, но и нерегулярность положения момента, соответствующего этому среднему значению тока, на временной оси, а следовательно, и относительную нестабильность частоты (Df/f) генерируемых коллективом пейсмейкеров колебаний
![]() .
.
Таким образом, увеличение количества пейсмейкеров стабилизирует частоту импульсов возбуждения, вырабатываемых центром автоматизма. В сердце человека имеется около 5000 клеток-генераторов. Следовательно, относительная нестабильность составит единицы процентов, что и наблюдается на практике.
 Сердце
имеет еще и нервную регуляцию частоты и силы мышечных сокращений с помощью так
называемых симпатической и парасимпатической нервных систем. Сигналы из первой
приводят к увеличению частоты и скорости проведения импульсов, второй – к противоположному
результату.
Сердце
имеет еще и нервную регуляцию частоты и силы мышечных сокращений с помощью так
называемых симпатической и парасимпатической нервных систем. Сигналы из первой
приводят к увеличению частоты и скорости проведения импульсов, второй – к противоположному
результату.
Сердце имеет три центра автоматизма (рис. 38). Основной из них – синоатриальный (СА) узел 1. Он генерирует импульсы с частотой 60-80 в минуту. Сигнал, вырабатываемый пейсмейкерами в СА узле, распространяется по специальной системе мышечных клеток, называемой проводящей. Особенностью этих клеток является большая скорость распространения возбуждения в сравнении со скоростью в сократительном миокарде. Потенциал действия, возникший в СА узле, передается на сократительные мышцы правого 7 и левого 2 предсердий и по трем веткам проводящей системы в правом предсердии в узел 3, находящийся на границе предсердий и желудочков. Этот узел называется атриовентрикулярным (АВ). Скорость распространения возбуждения по предсердным участкам проводящей системы составляет 30-80 см/с. В АВ узле происходит резкое уменьшение скорости распространения сигнала до 2-5 см/с. Этим обеспечивается необходимая последовательность возбуждения предсердий и желудочков. Из АВ узла возбуждение передается во внутрижелудочковую проводящую систему, содержащую пучок Гиса 4, оканчивающийся волокнами Пуркинье. Скорость проведения сигнала в пучке Гиса составляет 100-150 см/с, а в волокнах Пуркинье – 300-400 см/с. Такая дифференцировка скорости обеспечивает практически одновременный охват возбуждением желудочков и эффективный выброс крови. Область перехода АВ узла в пучок Гиса (АВ-соединение) является узлом автоматизма второго порядка. Пейсмейкеры этой активной области могут спонтанно возбуждаться с частотой 40-60 импульсов в минуту. Узлами автоматизма третьего порядка являются ножки пучка Гиса и волокна Пуркинье. Активность пейсмейкеров этих элементов проводящей системы обеспечивает формирование потенциала действия с частотой 25-45 импульсов в минуту. Как уже упоминалось, основным центром автоматизма является СА-узел. Если он функционирует, то остальные подавляются. Каждый из последующих центров проявляет свою активность только в случае прекращения функционирования центра более высокого порядка.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.