В состоянии покоя (![]() ) ток во внешней цепи (выходной цепи
усилителя и измерительного электрода) не течет, т.е. сумма токов ветвей равна
нулю. Токи отдельных ветвей, однако, могут быть отличны от нуля. В частности,
существование тока утечки через мембрану в состоянии покоя влечет за собой с
необходимостью существование тока еще хотя бы одного канала. При скачкообразном
уменьшении абсолютного значения потенциала мембраны (деполяризации) изменяются
вероятности открывания (а, следовательно, и проводимости) натриевых и калиевых
каналов. После подачи ступенчатого сигнала на вход
) ток во внешней цепи (выходной цепи
усилителя и измерительного электрода) не течет, т.е. сумма токов ветвей равна
нулю. Токи отдельных ветвей, однако, могут быть отличны от нуля. В частности,
существование тока утечки через мембрану в состоянии покоя влечет за собой с
необходимостью существование тока еще хотя бы одного канала. При скачкообразном
уменьшении абсолютного значения потенциала мембраны (деполяризации) изменяются
вероятности открывания (а, следовательно, и проводимости) натриевых и калиевых
каналов. После подачи ступенчатого сигнала на вход ![]() операционного
усилителя ОУ1 (рис. 9) напряжение на входе
операционного
усилителя ОУ1 (рис. 9) напряжение на входе ![]() стремится
к уровню
стремится
к уровню ![]() по закону
по закону 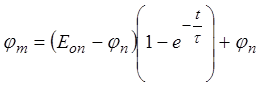 .
Постоянная времени t
определяется временем установления усилителя и постоянной времени емкости
мембраны, заряд которой происходит по цепи: выход усилителя ОУ1, токовый
электрод, измерительный электрод, общая шина (потенциалы инвертирующего и
неинвертирующего входов ОУ2 приблизительно равны). Через интервал времени (2-3)t конденсатор С полностью
заряжается, и напряжение на мембране достигает уровня
.
Постоянная времени t
определяется временем установления усилителя и постоянной времени емкости
мембраны, заряд которой происходит по цепи: выход усилителя ОУ1, токовый
электрод, измерительный электрод, общая шина (потенциалы инвертирующего и
неинвертирующего входов ОУ2 приблизительно равны). Через интервал времени (2-3)t конденсатор С полностью
заряжается, и напряжение на мембране достигает уровня ![]() .
Как уже упоминалось, при правильном выборе элементов схемы это время
значительно меньше длительности процессов активации натриевых и калиевых
каналов и, таким образом, изменение токов через эти каналы за время заряда
конденсатора оказывается несущественным. Процессы развития регенерации в
наибольшей степени проявляются после установления на мембране уровня напряжения
.
Как уже упоминалось, при правильном выборе элементов схемы это время
значительно меньше длительности процессов активации натриевых и калиевых
каналов и, таким образом, изменение токов через эти каналы за время заряда
конденсатора оказывается несущественным. Процессы развития регенерации в
наибольшей степени проявляются после установления на мембране уровня напряжения
![]() . Если
. Если ![]() меньше
порогового уровня возбуждения
меньше
порогового уровня возбуждения ![]() (
(![]() и
и ![]() отрицательны),
то изменения составляющих мембранного тока происходят в полном соответствии с
законами теории линейных электрических цепей: ток произвольной ветви
отрицательны),
то изменения составляющих мембранного тока происходят в полном соответствии с
законами теории линейных электрических цепей: ток произвольной ветви ![]() , содержащей источник
, содержащей источник ![]() и проводимости
и проводимости ![]() , определяется формулой
, определяется формулой ![]() . Общий ток во внешней цепи в
отсутствие возбуждения
. Общий ток во внешней цепи в
отсутствие возбуждения ![]() будет равен сумме токов ветвей.
В нашем случае
будет равен сумме токов ветвей.
В нашем случае ![]() . Если же
. Если же ![]() превышает пороговый уровень
возбуждения, то изменения натриевой и калиевой составляющих тока перестают
зависеть от потенциала мембраны линейным образом. Зависимость от времени общего
тока, регистрируемая во внешней цепи, при этом резко отличается от аналогичной
зависимости для случая отсутствия возбуждения. Во-первых, ток становится
двуполярным, и, во-вторых, значительно увеличивается его амплитуда. Используя
методы блокировки каналов, а также заменяя состав окружающей клетку среды,
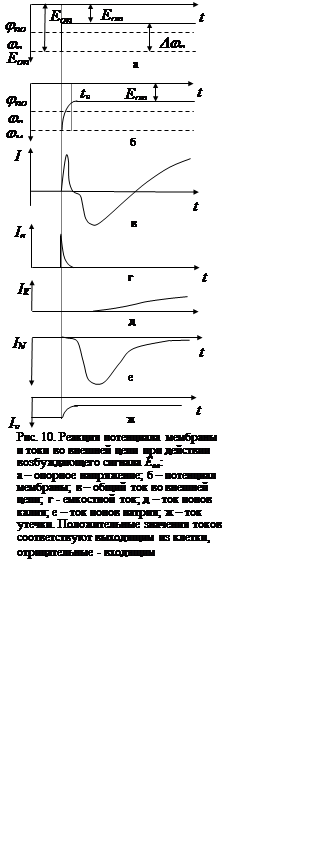
удалось осуществить разделение тока через мембрану на составляющие. Емкостной
ток (рис. 10, г), связанный с потенциалом мембраны соотношением
превышает пороговый уровень
возбуждения, то изменения натриевой и калиевой составляющих тока перестают
зависеть от потенциала мембраны линейным образом. Зависимость от времени общего
тока, регистрируемая во внешней цепи, при этом резко отличается от аналогичной
зависимости для случая отсутствия возбуждения. Во-первых, ток становится
двуполярным, и, во-вторых, значительно увеличивается его амплитуда. Используя
методы блокировки каналов, а также заменяя состав окружающей клетку среды,
удалось осуществить разделение тока через мембрану на составляющие. Емкостной
ток (рис. 10, г), связанный с потенциалом мембраны соотношением 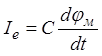 , протекает в течение времени
установления потенциала
, протекает в течение времени
установления потенциала ![]() (рис. 10,
б). Калиевый ток (рис. 10, д) монотонно нарастает во времени, стремясь к
определенному пределу, и держится на нем в течение достаточно длительного времени
(сотни миллисекунд). Натриевый ток (рис. 10, е) имеет колоколообразную форму.
Длительность его составляет единицы миллисекунд (2-3). Скорость нарастания
натриевого тока в начальной фазе возбуждения значительно превосходит скорость
нарастания калиевого тока. Вследствие этого в начале деполяризации общий ток
мембраны определяется натриевой компонентой и является втекающим. В дальнейшем
натриевый ток достигает максимума и начинает уменьшаться. К этому времени
возрастает калиевый ток, так что сумма токов становится положительной –
вытекающей и стремящейся к стационарному уровню калиевого тока. На баланс токов
некоторое влияние оказывает ток утечки (рис. 10, ж), изменяющийся с момента
подачи скачка опорного напряжения с одного стационарного уровня на другой по
закону изменения мембранного потенциала. Вклад в этот ток осуществляют
проводимости невозбудимых ионных каналов различной природы (
(рис. 10,
б). Калиевый ток (рис. 10, д) монотонно нарастает во времени, стремясь к
определенному пределу, и держится на нем в течение достаточно длительного времени
(сотни миллисекунд). Натриевый ток (рис. 10, е) имеет колоколообразную форму.
Длительность его составляет единицы миллисекунд (2-3). Скорость нарастания
натриевого тока в начальной фазе возбуждения значительно превосходит скорость
нарастания калиевого тока. Вследствие этого в начале деполяризации общий ток
мембраны определяется натриевой компонентой и является втекающим. В дальнейшем
натриевый ток достигает максимума и начинает уменьшаться. К этому времени
возрастает калиевый ток, так что сумма токов становится положительной –
вытекающей и стремящейся к стационарному уровню калиевого тока. На баланс токов
некоторое влияние оказывает ток утечки (рис. 10, ж), изменяющийся с момента
подачи скачка опорного напряжения с одного стационарного уровня на другой по
закону изменения мембранного потенциала. Вклад в этот ток осуществляют
проводимости невозбудимых ионных каналов различной природы (![]() ,
,![]() ).
Особенности поведения токов натриевых и калиевых каналов обусловлены изменением
их проводимостей. До поступления возбуждающего стимула калиевая и натриевая
проводимости остаются постоянными во времени и равны приблизительно 1 мС/см2
и 0,04 мС/см2 соответственно. На пике возбуждения натриевая
проводимость, как уже упоминалось, увеличивается в 500 раз и составляет 20
мС/см2, а калиевая в этот момент остается практически без изменения.
Существенный сдвиг калиевой проводимости наступает позже (через 3-5 мс).
).
Особенности поведения токов натриевых и калиевых каналов обусловлены изменением
их проводимостей. До поступления возбуждающего стимула калиевая и натриевая
проводимости остаются постоянными во времени и равны приблизительно 1 мС/см2
и 0,04 мС/см2 соответственно. На пике возбуждения натриевая
проводимость, как уже упоминалось, увеличивается в 500 раз и составляет 20
мС/см2, а калиевая в этот момент остается практически без изменения.
Существенный сдвиг калиевой проводимости наступает позже (через 3-5 мс).
 На рис. 11 приведены зависимости
упомянутых проводимостей мембраны аксона кальмара при различных значениях напряжения
на ней, полученные в экспериментах с фиксацией потенциала.
На рис. 11 приведены зависимости
упомянутых проводимостей мембраны аксона кальмара при различных значениях напряжения
на ней, полученные в экспериментах с фиксацией потенциала.
При деполяризации мембраны до
уровня, превышающего критический, калиевая проводимость, монотонно
увеличиваясь, достигает постоянного значения, определяемого мембранным потенциалом.
Чем больше этот потенциал, тем выше установившийся уровень, однако существует
максимальное значение проводимости ![]() , которое невозможно
превзойти ни при каких значениях потенциала. Натриевая проводимость растет
значительно быстрее и в течение короткого времени достигает максимума. Скорость
нарастания и величина максимума увеличиваются по мере роста мембранного
потенциала. Однако и здесь, как и в случае калиевых каналов, имеется предельный
уровень. Спад натриевой проводимости (инактивация) происходит также быстро.
Скорость его тем больше, чем больше деполяризация.
, которое невозможно
превзойти ни при каких значениях потенциала. Натриевая проводимость растет
значительно быстрее и в течение короткого времени достигает максимума. Скорость
нарастания и величина максимума увеличиваются по мере роста мембранного
потенциала. Однако и здесь, как и в случае калиевых каналов, имеется предельный
уровень. Спад натриевой проводимости (инактивация) происходит также быстро.
Скорость его тем больше, чем больше деполяризация.
 Характер
поведения натриевых и калиевых каналов объясняется их активацией в начальной
стадии возбуждения и инактивацией в конечной. Тонкая структура механизмов
активации и инактивации в настоящее время в достаточной мере не раскрыта.
Существует несколько моделей этих процессов. Одна из них предполагает наличие в
мембране неких подвижных частиц, перераспределяемых под действием электрического
поля и, таким образом, управляющих состоянием каналов. Другая – связывает
изменения проницаемостей со структурированием ориентации электрических диполей,
расположенных внутри каналов. Наиболее вероятна, однако, версия, основанная на
предположении о скачкообразных конформационных перестройках белковых молекул
мембраны (ответственных за ее проницаемость) под действием электрического поля.
От напряженности электрического поля в этом случае должны зависеть как
вероятность конформационного процесса, так и время жизни в модификации с
увеличенной проницаемостью канала. На основании многочисленных опытных данных
были сформулированы основные особенности транспорта ионов через возбудимые мембраны.
Характер
поведения натриевых и калиевых каналов объясняется их активацией в начальной
стадии возбуждения и инактивацией в конечной. Тонкая структура механизмов
активации и инактивации в настоящее время в достаточной мере не раскрыта.
Существует несколько моделей этих процессов. Одна из них предполагает наличие в
мембране неких подвижных частиц, перераспределяемых под действием электрического
поля и, таким образом, управляющих состоянием каналов. Другая – связывает
изменения проницаемостей со структурированием ориентации электрических диполей,
расположенных внутри каналов. Наиболее вероятна, однако, версия, основанная на
предположении о скачкообразных конформационных перестройках белковых молекул
мембраны (ответственных за ее проницаемость) под действием электрического поля.
От напряженности электрического поля в этом случае должны зависеть как
вероятность конформационного процесса, так и время жизни в модификации с
увеличенной проницаемостью канала. На основании многочисленных опытных данных
были сформулированы основные особенности транспорта ионов через возбудимые мембраны.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.