Здесь ![]() -
потенциал мембраны (общая шина – межклеточная жидкость), Е1, Е2,
… Еi – равновесные мембранные
потенциалы различных типов ионов (K+, Na+, Cl– и
т.д.), R1, R2,
… Ri – сопротивления ветвей
(каналов), I1, I2,
… Ii – токи ветвей (каналов).
-
потенциал мембраны (общая шина – межклеточная жидкость), Е1, Е2,
… Еi – равновесные мембранные
потенциалы различных типов ионов (K+, Na+, Cl– и
т.д.), R1, R2,
… Ri – сопротивления ветвей
(каналов), I1, I2,
… Ii – токи ветвей (каналов).
Потенциал ![]() в данном случае находится из условия
в данном случае находится из условия
![]() . Ток каждой ветви определяется
формулой
. Ток каждой ветви определяется
формулой 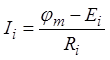 . Подставив полученное в предыдущую
формулу, приведем ее к виду
. Подставив полученное в предыдущую
формулу, приведем ее к виду 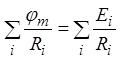 , из которого с
учетом
, из которого с
учетом 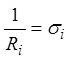 и следует соотношение (8).
Эквивалентная схема особенно убедительно свидетельствует о необходимости учета
ветвей с
и следует соотношение (8).
Эквивалентная схема особенно убедительно свидетельствует о необходимости учета
ветвей с ![]() (
(![]() )
при расчете мембранного потенциала.
)
при расчете мембранного потенциала.
Мембранный потенциал имеет большое
значение в жизнедеятельности организма. Существование его является одним из
факторов, обеспечивающих процессы метаболизма: за счет мембранного потенциала
осуществляется электрический транспорт различных веществ в клетку и из нее.
Мембранный потенциал обеспечивает также защиту клетки от воздействия внешних
электрических полей. Суть этой защиты сводится к тому, что значение
напряженности поля в мембране, обусловленное ее собственным потенциалом, во
много раз превосходит напряженности электрических полей стандартной среды
обитания на Земле. В самом деле, порядок напряженности поля внутри мембраны (а
именно напряженность является силовой функцией электрического поля, ![]() ) определяется как отношение
мембранного потенциала
) определяется как отношение
мембранного потенциала ![]() к толщине мембраны d:
к толщине мембраны d: ![]() .
.
Для характерных ![]() = 100 mB и d = 10 нм напряженность поля составляет 10 кВ/мм. Это
громадная величина. Она является предельной для вакуумных приборов, т.е. только
в вакууме могут быль реализованы столь большие поля. В обычной же среде
обитания человека (при атмосферном давлении 760 мм рт.ст., температуре ~ 20 °С) удельные напряженности не превышают 1
кВ/мм, т.е. имеют примерно на порядок меньшие значения, чем в мембране. Это
связано с тем обстоятельством, что в большинстве случаев область, в которой
создается электрическое поле (переменное или постоянное) превышает длину
свободного пробега в газе при давлении 760 мм рт.ст. и температуре 20 °С. Поэтому увеличение поля сверх 1 кВ/мм
приводит к пробою газа и шунтированию источника напряжения, формирующего поле.
И только в том случае, если протяженность пространства, в котором существует
поле, не превышает длины свободного пробега в газе, т.е. воздух при давлении
760 мм рт.ст. удовлетворяет определению понятия вакуума, пробой может не
наступить. Длина свободного пробега в воздухе
в стандартных условиях имеет порядок l=10-7–10-8
м =10-100 нм. Чтобы протяженность области поля с напряженностью до 10 кВ/мм не
превышала l, необходимо, чтобы это поле было
переменным и имело длину волны l
порядка l, т.е. l » 10-100 нм. Такую
длину волны имеют рентгеновские лучи и примыкающие к ним электромагнитные волны
в области вакуумного ультрафиолета. Излучение с этими и меньшими длинами волн
могут оказать существенное влияние на процессы на мембране клетки и в ее объеме.
= 100 mB и d = 10 нм напряженность поля составляет 10 кВ/мм. Это
громадная величина. Она является предельной для вакуумных приборов, т.е. только
в вакууме могут быль реализованы столь большие поля. В обычной же среде
обитания человека (при атмосферном давлении 760 мм рт.ст., температуре ~ 20 °С) удельные напряженности не превышают 1
кВ/мм, т.е. имеют примерно на порядок меньшие значения, чем в мембране. Это
связано с тем обстоятельством, что в большинстве случаев область, в которой
создается электрическое поле (переменное или постоянное) превышает длину
свободного пробега в газе при давлении 760 мм рт.ст. и температуре 20 °С. Поэтому увеличение поля сверх 1 кВ/мм
приводит к пробою газа и шунтированию источника напряжения, формирующего поле.
И только в том случае, если протяженность пространства, в котором существует
поле, не превышает длины свободного пробега в газе, т.е. воздух при давлении
760 мм рт.ст. удовлетворяет определению понятия вакуума, пробой может не
наступить. Длина свободного пробега в воздухе
в стандартных условиях имеет порядок l=10-7–10-8
м =10-100 нм. Чтобы протяженность области поля с напряженностью до 10 кВ/мм не
превышала l, необходимо, чтобы это поле было
переменным и имело длину волны l
порядка l, т.е. l » 10-100 нм. Такую
длину волны имеют рентгеновские лучи и примыкающие к ним электромагнитные волны
в области вакуумного ультрафиолета. Излучение с этими и меньшими длинами волн
могут оказать существенное влияние на процессы на мембране клетки и в ее объеме.
Таким образом, мембранный потенциал имеет чрезвычайно важное значение для существования организма.
1.3. Возбуждение мембраны. Потенциал действия
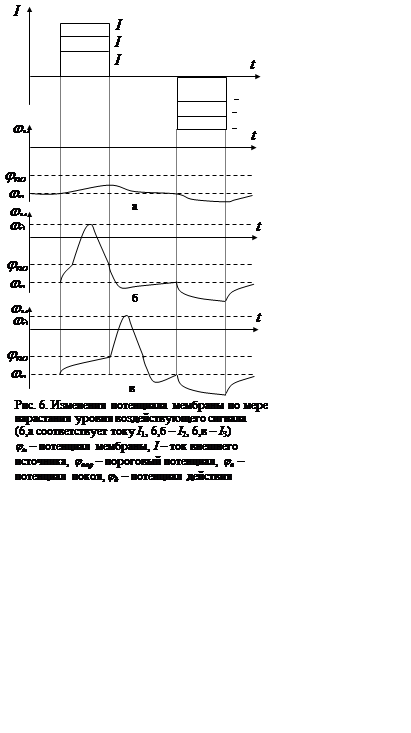
Нервные и мышечные клетки обладают особенностью, которая отличает их от клеток других органов и тканей. Особенность эта заключается в том, что потенциал их мембраны перестает линейно зависеть от воздействия при превышении последним определенного уровня. Параметры импульса напряжения на мембране (амплитуда и длительность) при этом определяются лишь характеристиками мембраны и окружающих ее электролитов. Процесс изменения напряжения на мембране клетки, характеризующийся нелинейными и параметрическими проявлениями в ответ на внешнее воздействие, называется возбуждением. Максимальное напряжение на мембране, возникающее при ее возбуждении внешним сигналом, превышающим пороговый уровень, называется потенциалом действия (ПД). Механизм возбуждения нервных и мышечных клеток лежит в основе информационной системы организма. Полагают, что упомянутые клетки в процессе эволюции сменили некоторые функции, связанные с транспортом веществ через мембрану, на функции, обеспечивающие информационный обмен между различными структурами. Качественно картина изменения потенциала мембраны нервной клетки при различных уровнях воздействия представлена на рис. 6 (потенциал отсчитывается от внешней поверхности мембраны, точнее, от точки в объеме межклеточной жидкости, близкой к поверхности мембраны).
В покое мембрана имеет потенциал ![]() , определяемый формулой (8) (в данном
случае мы заменили разность потенциалов
, определяемый формулой (8) (в данном
случае мы заменили разность потенциалов ![]() с
неопределенной точкой отсчета потенциалом, отсчитываемым от внешней поверхности
мембраны). При анализе переходных процессов под воздействием внешнего источника
электрического тока в эквивалентной схеме мембраны должнабыть учтена ее емкость
С. Заменив суммарное действие всех источников
с
неопределенной точкой отсчета потенциалом, отсчитываемым от внешней поверхности
мембраны). При анализе переходных процессов под воздействием внешнего источника
электрического тока в эквивалентной схеме мембраны должнабыть учтена ее емкость
С. Заменив суммарное действие всех источников ![]() с
проводимостями
с
проводимостями ![]() (рис. 5) одним источником
(рис. 5) одним источником ![]() с проводимостью 1/R, эквивалентную схему для качественного рассмотрения
изменений мембранного потенциала при внешнем воздействии можно представить в
виде, изображенном на рис. 7.
с проводимостью 1/R, эквивалентную схему для качественного рассмотрения
изменений мембранного потенциала при внешнем воздействии можно представить в
виде, изображенном на рис. 7.
 При подключении мембраны к цепи
внешнего источника питания I(t) потенциал ее j
начинает изменяться по закону, определяемому дифференциальным уравнением
При подключении мембраны к цепи
внешнего источника питания I(t) потенциал ее j
начинает изменяться по закону, определяемому дифференциальным уравнением 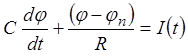 . Учитывая, что в момент подключения
источника тока потенциал мембраны был равен потенциалу покоя
. Учитывая, что в момент подключения
источника тока потенциал мембраны был равен потенциалу покоя ![]() , и полагая, что I(t) представляет собой импульсный ток прямоугольной формы
амплитудой
, и полагая, что I(t) представляет собой импульсный ток прямоугольной формы
амплитудой ![]() и длительностью
и длительностью ![]() , решение уравнения можно представить
следующим образом:
, решение уравнения можно представить
следующим образом:
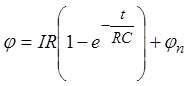 . (16)
. (16)
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.