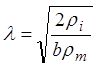 .
.
Зададимся
численными значениями: ![]() = 104 Ом×см2 = 1 Ом×м2;
= 104 Ом×см2 = 1 Ом×м2;
![]() = 102 Ом×см
= 1 Ом×м; b =
0,2 мм = 2×10-4 м. Подставив
их в последнюю формулу, приходим к значению l
= 1 1/см. Величина 1/l характеризует
расстояние, на котором сигнал ослабляется в «е» раз. Хотя 1/l и мало в сравнении с размерами позвоночных,
для мелких существ, например насекомых, этого расстояния вполне достаточно для
передачи сигнала без дополнительных затрат энергии.
= 102 Ом×см
= 1 Ом×м; b =
0,2 мм = 2×10-4 м. Подставив
их в последнюю формулу, приходим к значению l
= 1 1/см. Величина 1/l характеризует
расстояние, на котором сигнал ослабляется в «е» раз. Хотя 1/l и мало в сравнении с размерами позвоночных,
для мелких существ, например насекомых, этого расстояния вполне достаточно для
передачи сигнала без дополнительных затрат энергии.
Качественно характер зависимости l от параметров линии передачи (R, G) можно объяснить тем, что каждый элементарный отрезок линии представляет собой делитель из соединенных последовательно резисторов R и 1/G. Напряжение, снимаемое с G, тем больше, чем меньше R и G и, следовательно, тем больше расстояние, на которое распространяется заданный уровень.
Оценим частоту сигнала, на которой постоянная затухания увеличивается в два раза в сравнении с постоянной затухания при w = 0
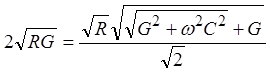 .
.
После
преобразований частота ![]() , соответствующая двукратному
увеличению l, определяется
соотношением
, соответствующая двукратному
увеличению l, определяется
соотношением ![]() , что приводит после выражения
G и С через удельные параметры к
, что приводит после выражения
G и С через удельные параметры к ![]() . Принимая
. Принимая ![]() = 1 мкФ/см2
= 10-2 Ф/м,
= 1 мкФ/см2
= 10-2 Ф/м, ![]() = 1 Ом×м2, получаем
= 1 Ом×м2, получаем ![]() Гц.
Гц.
2. Определим скорость распространения сигнала по нервному волокну.
Фазовая скорость ![]() определяется как скорость
поверхности постоянной фазы, т.е.
определяется как скорость
поверхности постоянной фазы, т.е. 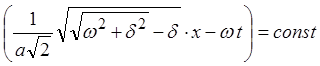 ;
;
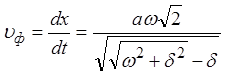 .
.
Отсюда следует,
что нервное волокно характеризуется сильной дисперсией: ![]() .
Таким образом, сложный сигнал, представляемый суммой гармонических
составляющих, будет искажаться по мере распространения. В области низких
частот, определяемых в данном случае неравенством w << d
с учетом
.
Таким образом, сложный сигнал, представляемый суммой гармонических
составляющих, будет искажаться по мере распространения. В области низких
частот, определяемых в данном случае неравенством w << d
с учетом 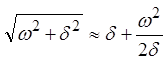 , фазовая скорость перестает зависеть
от частоты
, фазовая скорость перестает зависеть
от частоты
![]() .
.
Подстановка
значений a и d
дает ![]() .
.
Отсутствие
дисперсии означает, что в области низких частот групповая скорость ![]() , т.е. скорость переноса энергии,
также равна
, т.е. скорость переноса энергии,
также равна ![]() .
.
В общем случае групповая скорость может быть определена из соотношения
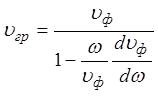 .
.
Записав 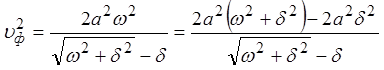 , используя
, используя 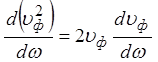 и
и
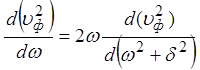 , можно прийти к выражению:
, можно прийти к выражению:
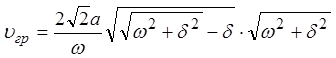 .
.
В области низких частот 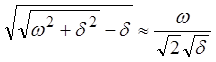 групповая скорость перестает
зависеть от частоты и совпадает (как и должно быть) с фазовой.
групповая скорость перестает
зависеть от частоты и совпадает (как и должно быть) с фазовой.
При w ® ¥, ![]() ® ¥,
что означает несправедливость формулы в области высоких частот. В этом случае
скорость должна определяться из решения задачи в электродинамической
постановке.
® ¥,
что означает несправедливость формулы в области высоких частот. В этом случае
скорость должна определяться из решения задачи в электродинамической
постановке.
Оценим величину скорости
распространения сигнала в пассивном волокне. Для конкретного примера выберем
значения b, ![]() ,
, ![]() такими же, как и в случае вычисления
l, а значение удельной емкости
такими же, как и в случае вычисления
l, а значение удельной емкости ![]() примем равным 1 мкФ/см2.
Формула для скорости, выраженная через удельные параметры волокна, примет вид:
примем равным 1 мкФ/см2.
Формула для скорости, выраженная через удельные параметры волокна, примет вид:
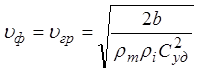 .
.
После подстановки численных
значений получаем ![]() м/с. Для биологических
объектов, имеющих размеры около сантиметра, этой скорости вполне достаточно:
время распространения сигнала имеет порядок единиц миллисекунд, что обеспечивает
требуемую быстроту реакции. На малых расстояниях несущественна также дисперсия:
разница в фазовых скоростях гармоник сигнала в этом случае еще не приводит к
сильному искажению волнового пакета.
м/с. Для биологических
объектов, имеющих размеры около сантиметра, этой скорости вполне достаточно:
время распространения сигнала имеет порядок единиц миллисекунд, что обеспечивает
требуемую быстроту реакции. На малых расстояниях несущественна также дисперсия:
разница в фазовых скоростях гармоник сигнала в этом случае еще не приводит к
сильному искажению волнового пакета.
Качественно характер зависимости скорости от параметров линии может быть объяснен тем, что постоянная времени цепи заряда и разряда конденсатора С определяется параллельными R и 1/G, т.е. отношением R/G. Чем меньше это отношение и емкость конденсатора С, тем быстрее осуществляются его заряд и разряд, а следовательно, увеличивается скорость передачи возмущения.
Из формулы для скорости видно, что она может быть увеличена, при прочих равных условиях, путем увеличения диаметра волокна. Однако этот путь приводит к тому, что нервные волокна (а их для анализа жизненной ситуации и реакции на нее необходимо достаточно много) начинают занимать значительное место в организме. Поэтому большой диаметр имеют, как правило, только волокна, наиболее важные для жизнедеятельности.
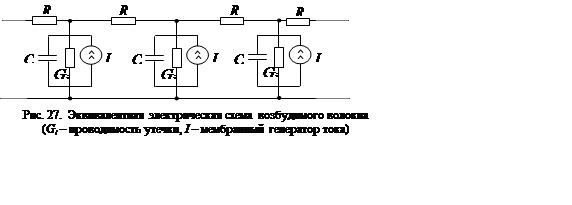
Механизм распространения сигнала в возбудимых нервных волокнах при уровнях, превышающих пороговый, существенно отличается от механизма передачи сигнала по линиям с проводниками, выполненными из металла. Если в последних ток практически одинаков во всех сечениях (при длине линии меньшей четверти длины волны), то в первых он неодинаков и отличен от нуля лишь в локальной области, перемещающейся по волокну. Распространение сигнала в этом случае иллюстрируется рис. 26. На нем изображено распределение зарядов и потенциала мембраны аксона для нескольких моментов времени.
Из рисунка видно, что сигнал по нервному волокну распространяется в виде локализованного в пространстве возмущенного участка. Токи через мембрану текут только в пределах этого участка. Слева к области возбуждения примыкает область в состоянии рефрактерности (через нее импульс возбуждения уже прошел), а справа – невозмущенная область. Вследствие этого сигнал распространяется только слева направо, захватывая невозмущенные участки по мере увеличения потенциала на фронте возмущенных до порогового уровня. Особенностью распространяющегося сигнала является постоянство его ампли-туды, определяемой потенциалом действия данного типа нервного волокна (например, в случае аксона кальмара натриевым потенциалом).
Количественный анализ процесса
распространения импульса возбуждения по нервному волокну наиболее полно может
быть осуществлен на основании уравнений Ходжкина-Хаксли. Однако эта система не
допускает относительно простых аналитических решений. Вследствие этого
используют различные упрощающие приемы. В частности, полученное ранее уравнение
для пассивного волокна может быть распространено и на возбудимое, если учесть
зависимость проводимости мембраны G от времени и
потенциала. На практике, однако, заменяют произведение GU
на эквивалентный генератор тока. С учетом проводимости утечки ![]() расчетная эквивалентная
электрическая схема аксона приобретает вид, показанный на рис. 27.
расчетная эквивалентная
электрическая схема аксона приобретает вид, показанный на рис. 27.

 |
Уравнение для потенциала мембраны U рассматриваемой канализирующей системы может быть записано следующим образом:
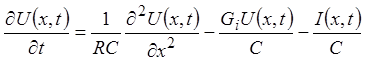 .
(25)
.
(25)
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.