![]()
Ветровое воздействие приводит к возникновению в приповерхностных горизонтах активных гидродинамических процессов, в результате чего рыба уходит в более глубокие слои воды (Герасимов, Поддубный, 1999; Малинин, Литвинов, 1991). Подобная реакция была зафиксирована во время ихтиологической съемки 1998 г. (табл. 5.6). Длительное ветровое воздействие вызвало миграцию рыб с мелководных местообитаний на смежные биотопы с большими глубинами и, в первую очередь, на затопленные русла рек. Русловые скопления рыб отличаются большой временной и пространственной стабильностью, однако их качественный и количественный состав существенно изменяется во времени в результате периодических, вызванных неблагоприятными воздействиями, миграций рыб с пойменных местообитаний. Этим объясняется более высокое видовое разнообразие уловов 1998 г. на русловых участках и сравнительно высокая доля в них плотвы и неполовозрелого леща, основные места нагула которых находятся на пойменных участках. При гидроакустической съемке на таких участках поймы регистрируется крайне низкая численность рыб (рис. 5.12 а). В это же самое время на прирусловой пойме, защищенной от ветрового воздействия, где волнение при сходной силе ветра гораздо ниже, чем в открытой части водохранилища, рыба не покидает пойму, образуя сравнительно плотные скопления (см. рис.
5.12 б).
Таблица 5.6
Ихтиомасса (кг/га) на двух русловых биотопах до шторма (над чертой) и сразу после шторма (под чертой) в августе-сентябре 1998 г.
|
Станции |
Размерные группы |
Общая ихтиомасса |
||
|
мелкая |
средняя |
крупная |
||
|
Бабьи горы |
|
145.7 162.0 |
|
|
|
Горькая соль |
70.9 110.9 |
323.0 584.6 |
371.9 |
1067.5 |

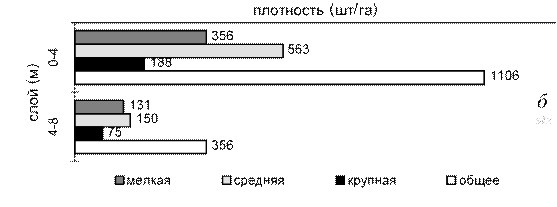
Рис. 5.12. Вертикальное распределение рыб разных размерных групп на пойменных местообитаниях открытого (а) и защищенного от ветра (б) участков водохранилища
На русловых местообитаниях при усилении ветра рыбы совершают вертикальные миграции, опускаясь в слои воды, недоступные для волнового воздействия, где образуют плотные скопления (рис. 5.13 б). На русловых участках, защищенных от ветрового воздействия (см. рис. 5.13 а), и по всей акватории водохранилища в штилевую погоду, как это происходилов 1999 г., рыбы формируют наиболее плотные скопления на глубине 4– 8 м (см. рис. 5.13 в). Следовательно, высокие значения ихтиомассы, зарегистрированные в 1998 г. на русловых станциях, являются следствием миграции рыб с пойменных местообитаний, подвергающихся сильному гидродинамическому воздействию.
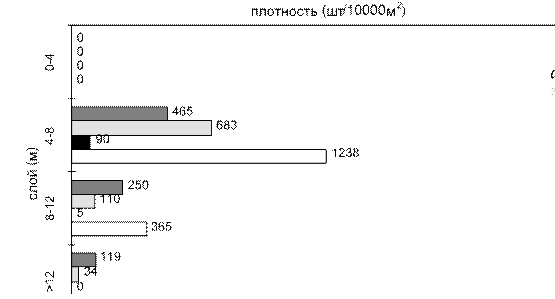

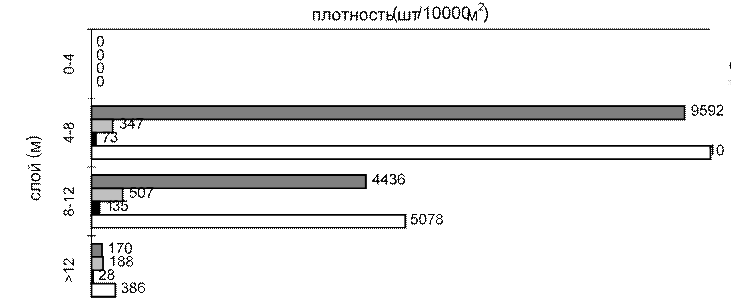
Рис. 5.13. Вертикальное распределение рыб:
а - русло на защищенном от ветра участке, б - в тот же день в открытой части, в - та же станция в открытой части, в штиль
Во время ихтиологической съемки 1999 г. общая ихтиомасса, а также ихтиомасса средней и крупной размерных групп на русловых местообитаниях была ниже (табл. 5.7), чем в 1998 г. Во время съемки скорость ветра не превышала 2–3 м/с. Распределение рыб по акватории было более равномерным (рис. 5.14), чем в 1998 г. Основное отличие заключалось в значительном увеличении ихтиомассы мелких рыб в 1999 г., обусловленном особенностями уровенного режима водохранилища (см. табл. 5.7). Чрезмерное для конца летнего сезона снижение уровня вызвало массовый выход молоди карповых рыб с прибрежных местообитаний на участки с большими глубинами, где они образовали плотные скопления. Основу этих скоплений составляла молодь плотвы и леща (см. табл. 5.4), что не характерно для русловых пелагических биотопов Рыбинского водохранилища.
Таблица 5.7
Средняя плотность рыб (M, экз./100 м2) в дневное и ночное время
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.