Ацилкарнитин обладает способностью проходить через внутреннюю мембрану митохондрий. На внутренней поверхности внутренней мембраны митохондрий ацилкарнитин взаимодействует с митохондриальным КоА. В результате в митохондриальном матриксе вновь образуется ацил-КоА, а карнитин высвобожда
Далее митохондриальный ацил-КоА распадается в результате повторяющейся последовательности из четырех реакций окисления с участием флавинадениндинуклеотида (ФАД), гидратации, окисления с участием НАД и тиолиза с участием КоА.
Краткая характеристика ферментов, катализирующих эти реакции:
Ацил~КоА дегидрогеназа. Есть несколько форм этого фермента, каждый из которых специфичен к ацил-КоА с определенной длиной углеводородной цепи. В ходе реакции, катализируемой этим ферментом, отщепляются два атома водорода, в результате образуется еноил-КоА с двойнойсвязью в транс-положении между a и b - углеродными атомами (С2 и С3). Образующийся ФАДН2 впоследствии окисляется в цепи тканевого дыхания с выделением энергии, которая запасается в двух молекулах АТФ.
-Еноил~КоА гидратаза. Фермент катализирует последующую реакцию гидратации по месту двойной связи с образованием 3-L-гидроксиацил~КоА. Гидроксильная группа в составе этого соединения находится у b-углеродного атома.
-b-оксиацил~КоА дегидрогеназа. Катализирует III этап - дегидрирование по месту b-углеродного атома с образованием кетогруппы и НАДН. Последний, окисляясь в цепи тканевого дыхания, приводит к высвобождению энергии, которая запасается в трех молекулах АТФ.
-Тиолаза. Катализирует четвертый этап, в ходе которого от b-кетоацил~КоА отщепляется двухуглеродный фрагмент в виде ацетил~КоА. Оставшийся ацил~КоА укорочен на 2 углеродных атома. Он опять включается в процесс (виток) b-окисления. Образовавшийся ацетил~КоА может поступать в цикл трикарбоновых кислот и, окисляясь там, давать энергию.
Цепь жирной кислоты укорачивается при этом на два атома углерода. Одновременно происходит образование ФАДН2, НАДН и ацетил-КоА. Укороченный ацил-КоА вступает вновь в следующий цикл окисления, начиная с реакции, катализируемой ацил-КоА-дегидрогеназой. Большой вклад в изучение этой последовательности реакций внесли Дэвид Грин, Северо Очоа и Феодор Линен. Они и назвали этот процесс b-окислением в соответствии с тем, что окислению всегда подвергается b-углеродный атом остатка жирной кислоты (второй углеродный атом ацила).
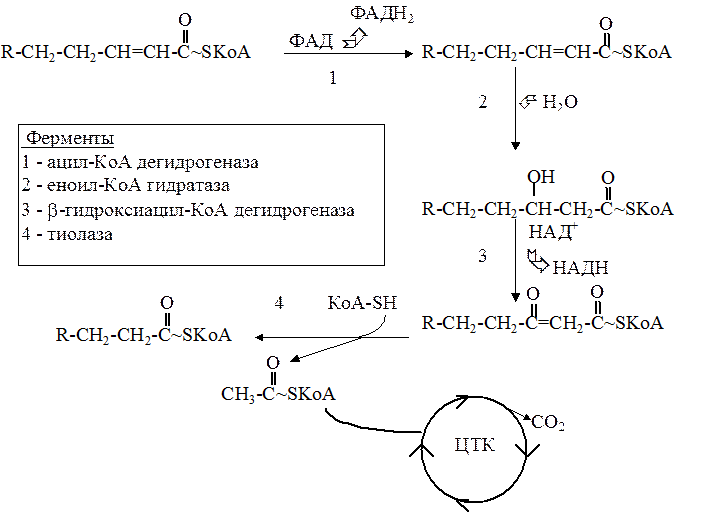
Последовательность реакций b-окисления жирных кислот: окисление, гидратация, окисление и тиолиз.
Конкуренция глюкозы и жирных кислот за использование в качестве субстратов: цикл Рэндэла. Увеличенное окисление жирных кислот ингибирует окисление глюкозы в клетках скелетных мышц и сердца за счет ингибирования пируватдегидрогеназы (соотношение ацетил~КоА/КоА-SH). При голодании такое явление призвано уменьшить утилизацию глюкозы периферическими тканями. Однако у людей с высоким уровнем СЖК это является одной из причин устойчивости к действию инсулина (к примеру, при диабете, беременности). С другой стороны, увеличение окисления глюкозы может ингибировать окисление жирных кислот. Это обусловлено тем, что регуляция поглощения жирных кислот митохондриями преимущественно осуществляется за счет контроля КПТI со стороны малонил-КоА, который выполняет роль аллостерического ингибитора этого фермента. Малонил-КоА - это начальный промежуточный продукт в синтезе жирных кислот, образованный из ацетил-КоА в цитоплазме. Избыток ацетил-КоА в митохондриях не может самостоятельно пройти в цитоплазму. Проход через митохондриальную мембрану становится возможным благодаря цитратному шунту. Ацетил-КоА карбоксилаза катализирует образование малонил-КоА. На эту реакцию расходуется СО2 и АТФ. Таким образом, условия, которые способствуют липогенезу (наличие большого количества глюкозы), подавляют b-окисление жирных кислот.
Гипергликемия частично подавляет липолиз. Энергетический выход окисления жирных кислот зависит от длины цепи.
Можно подсчитать энергетический выход b-окисления жирных кислот. В каждом цикле реакций ацил-КоА укорачивается на 2 углерода и образуется по одной молекуле ФАДН2, НАДН.Н+ и ацетил-КоА.
При окислении каждого из этих НАДН через дыхательную цепь образуется три молекулы АТФ, тогда как при окислении каждого ФАДН2 - две молекулы АТФ, потому что в этом случае электроны поступают в цепь на уровне кофермента Q (см. "тканевое дыхание"). Напомним, что окисление ацетил-КоА в цикле трикарбоновых кислот дает 12 молекул АТФ. Таким образом, энергетический выход 1 цикла b -окисления составляет 5 молекул АТФ + 12 молекул АТФ. Для подсчета энергетического выхода b-окисления конкретной жирной кислоты с четным числом углеродных атомов необходимо знать количество циклов b-окисления (оно составляет n/2 - 1, где n - число углеродных атомов в составе жирной кислоты) и молекул образующихся ацетил-КоА (оно составляет n/2). Из общей суммы АТФ необходимо вычесть одну молекулу АТФ, которая была затрачена на активацию жирной кислоты в начале всего процесса.
Реакции b-окисления тесно сопряжены друг с другом. Промежуточные продукты неизбежно переходят из одной реакции в другую; кроме наличия субстратов других контролирующих механизмов для этих реакций нет. Уровень b-окисления может возрастать при механической мышечной работе, при уменьшении соотношения ацетил-КоА/ацил-КоА, НАДН/НАД+ и ФАДН2/ФАД.
Энергетический выход b-окисления на примере пальмитиновой кислоты. Образование АТФ (2 АТФ/ФАДН2; 3 АТФ/НАДН; 12 АТФ/ацетил~КоА; таким образом для пальмитоил~КоА (жирная кислота с 16 С): 7 ФАДН2, 7 НАДН и 8 ацетил-КоА = 131 АТФ.
Расход АТФ на активацию - 1 АТФ (используется энергия гидролиза двух макроэргических связей), в ходе которой пальмитат превращается в пальмитоил-КоА. Таким образом, чистый энергетический выход для окисления пальмитата равен 130 АТФ.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.