
Олеиновая кислота (18:1 w 9) или цис - 9 - октадеценовая кислота.
Линолевая кислота (18:2 w 6) или цис -9-цис-12 -октадекадиеновая кислота.
Линоленовая кислота (18:3 w 3) или цис-9, цис-12, цис-15-октадека-триеновая кислота.
Арахидоновая кислота (20:4 w 6) или (эйкозатетраен-5,8,11,14-овая кислота), цис-5, цис-8, цис-11, цис-14- эйкозатетраеновая кислота.
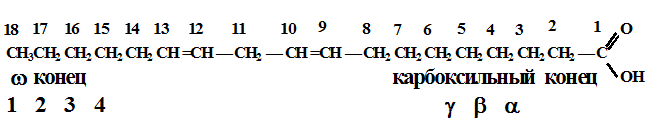
Характерно, что все двойные связи в составе жирных кислот организма имеют цис-конфигурацию Двумя преобладающими мононенасыщенными жирными кислотами животных липидов являются олеиновая и пальмитолеиновая. Олеиновая кислота наиболее широко распространена в природе и преобладает в количественном отношении. Среди полиненасыщенных жирных кислот в тканях млекопитающих наиболее часто встречается линолевая кислота, содержащая две двойные связи, линоленовая - с тремя двойными связями и арахидоновая - с четырьмя двойными связями. Ненасыщенность жирных кислот существенно влияет на их свойства. С увеличением числа двойных связей снижается температура плавления жирных кислот, возрастает их растворимость в неполярных растворителях. Все ненасыщенные жирные кислоты, встречающиеся в природе, при комнатной температуре являются жидкостями. Олеиновая кислота (18:1 w 9) или цис - 9 - октадеценовая кислота.
Линолевая кислота (18:2 w 6) или цис -9-цис-12 -октадекадиеновая кислота.
Линоленовая кислота (18:3 w 3) или цис-9, цис-12, цис-15-октадека-триеновая кислота.
Арахидоновая кислота (20:4 w 6) или (эйкозатетраен-5,8,11,14-овая кислота), цис-5, цис-8, цис-11, цис-14- эйкозатетраеновая кислота.
Транспорт свободных жирных кислот (СЖК) в плазме крови
Как уже упоминалось, основными потребителями СЖК являются клетки сердечной и скелетной мышц, печени. Для этих тканей характерно поглощение 30%-40% СЖК, содержащихся в артериальной крови. Основными СЖК плазмы крови являются пальмитиновая и олеиновая.
Комплекс СЖК - альбумин. Вследствие сильной гидрофобности >99% жирных кислот плазмы циркулируют в нековалетно связанном с альбумином состоянии. Важнейшей функцией альбумина, наиболее распространенного белка во внеклеточной жидкости, является связывание небольших, плохо растворимых в воде молекул. На молекуле альбумина имеется 3 центра связывания, к которым присоединяется 10 и более молекул жирных кислот. В плазме имеется небольшая концентрация жирных кислот, которые не связаны с альбумином, но уравновешены с комплексом альбумин-жирные кислоты. Они могут диффундировать через эндотелий капилляров и поглощаться тканями. Присутствие альбумина обеспечивает существование мощного резервуара жирных кислот в непосредственной близости к тканям всего организма.
Вклад СЖК плазмы крови в энергетический обмен. Период полужизни СЖК плазмы короток (около 1,5 мин), а оборот СЖК плазмы значительный и составляет около 200г/сутки. Если бы все они подверглись окислению, то выделилось бы 1800 ккал энергии. На самом деле, значительная часть этих жирных кислот используется для эстерификации ТАГ.
Оценка вклада СЖК плазмы крови в энергетический обмен
человека
|
Параметр |
Значение |
|
Масса |
70 кг |
|
Объем плазмы |
3 л |
|
СЖК плазмы (средн. концентрация) |
0,5 ммоль |
|
Молекулярная масса СЖК (средн.) |
280 |
|
Период полужизни СЖК плазмы |
1,5 мин |
|
Оборот СЖК плазмы |
10 ммоль/л 720 ммоль/сутки 201,6 г/сутки |
|
Калорический коэффициент оборота (9 ккал/г) |
1818 ккал/сутки |
|
Калорическая потребность (умеренная активность) |
2350 ккал/сут |
|
Максимальный вклад оборота СЖК плазмы в калорическую потребность |
77% |
Поглощение жирных кислот и роль белка, связывающего жирные кислоты. Считается, что жирные кислоты проходят через клеточную мембрану путем диффузии по концентрационному градиенту, хотя вопрос о том, существует ли такой градиент для жирных кислот, остается спорным. В последнее десятилетие получено много сведений относительно структуры и функции семейства белков, связывающих жирные кислоты. Эти белки обладают тканевой специфичностью и составляют от 2% до 5% от общего количества цитозольного белка в сердечной и скелетной мышцах, молочной железе, надпочечниках и мозге. Их роль заключается в обеспечении поглощения и внутриклеточного транспорта жирных кислот, в регуляции клеточного содержания жирных кислот и ацил~КоА. Высокая концентрация таких белков позволяет клеткам поддерживать внутриклеточный фонд жирных кислот, сохранять на низком уровне концентрацию несвязанных жирных кислот. Последние обладают детергентным действием.
Бета – окисление жирных кислот
Начинается с реакции RCOOH + HS~KoA + ATF ------- RCOO~SKoA + AMF + дифосфат
Реакция эта протекает, главным образом, в цитоплазме, в то время как процесс b-окисления жирных кислот происходит в митохондриях. Ацил-КоА не может проникнуть в митохондрию без помощи карнитина. Карнитин является широко распространенным соединением, особенно много его в мышцах. Образуется он из аминокислот лизина и метионина в печени и почках. На наружной стороне внутренней мембраны митохондрий имеется фермент ацилкарнитин трансфераза, который катализирует взаимодействие ацил-КоА с карнитином:
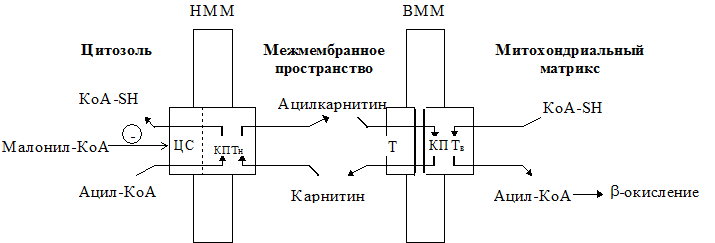
Опосредованный карнитином перенос длинноцепочечного ацил-КоА в митохондриальный матрикс
КПТн катализирует образование ацилкарнитинового комплекса из ацил-КоА и карнитина на внутренней стороне наружной митохондриальной мембраны (НММ). Ацилкарнитиновый комплекс затем диффундирует через межмембранное пространство к внутренней митохондриальной мембране (ВММ). Там совместное последовательное действие карнитин:ацилкарнитин транслоказы (Т) и КПТв обеспечивает поступление ацил-КоА в митохондриальный матрикс для последующего окисления. Активность КПТн ингибируется малонил-КоА на наружной стороне наружной мембраны митохондрий. Наличие специального места связывания малонил-КоА пока четко не установлено.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.