Таким образом, общая динамика катаболизма ЛПОНП заключается в потере ТАГ и приобретении ЭХ. Метаболизм ЛПОНП тесно связан с обменом ЛПВП и липопротеинлипазой.
Липопротеины промежуточной плотности (ЛППП). Из ЛПОНП в ходе описанных выше превращений образуются ЛППП. ЛППП не могут эффективно связываться с ЛПЛ, соответственно обмен ЛППП происходит с помощью другого липолитического фермента, печеночной липазы (ПЛ)[6]. Этот фермент синтезируется в гепатоцитах. В синтезе его также принимает участие синусоидальный эндотелий. ПЛ более эффективно, чем ЛПЛ, катализирует гидролиз ФЛ.
Вопрос 3. Связанная с рецептором частица ЛПНП подвергается поглощению клеткой Внутри образовавшихся эндосом липопротеины отщепляются от рецепторов. В дальнейшем ЛПНП поступают в лизосомы, где они разрушаются. В лизосомах происходит гидролиз эфиров холестерола, находившихся в составе ЛПНП. В результате образуется свободный холестерол или окисленные его формы. Свободный холестерол используется для различных целей. Он служит структурным компонентом клеточных мембран, субстратом для синтеза стероидных гормонов и желчных кислот. Продукты же его окислительного превращения оказывают регуляторное воздействие на организм.
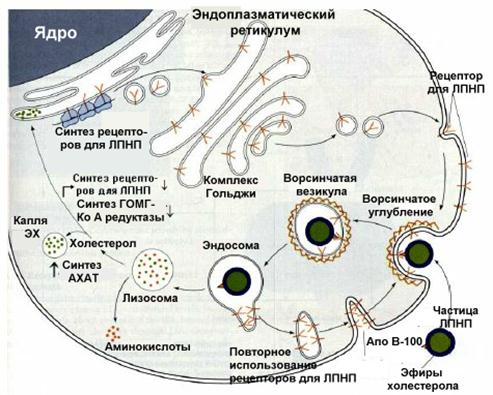
. Схема поступления в клетки ЛПНП
Контролирующие механизмы координируют использование внутри- и внеклеточных источников холестерола. При достаточном количестве ЛПНП клетки млекопитающих, с помощью рецепторов, преимущественно используют их в качестве источника холестерола. В это время внутриклеточная система синтеза холестерола находится как бы в резерве, не функционирует в полную силу. Дело в том, что внутриклеточное высвобождение из ЛПНП СХ или окисленных его форм, происходящее таким путем, воздействует на ряд процессов по принципу обратной связи и защищают клетку от чрезмерного накопления ХС. Во-первых, уменьшается активность 3-гидрокси-3-метилглютарил-КоА (ГОМГ-КоА) синтетазы и ГОМГ-КоА редуктазы, двух ключевых ферментов внутриклеточного биосинтеза ХС (см. раздел "Биосинтез ХС"). Во-вторых, стеролы активируют фермент ацил-КоА-холестерол ацил трансферазу, который катализирует этерификацию ХС. Это позволяет клеткам депонировать избыток ХС в форме своеобразных внутриклеточных капель, содержащих гидрофобные ЭХ. В-третьих, уменьшается синтез новых рецепторов к ЛПНП[7], что снижает поступление в клетку этих липопротеиновых частиц.
Обратный транспорт холестерола из периферических тканей к печени осуществляется посредством ЛПВП. Эти липопротеиновые частицы удаляют избыток свободного (неэстерифицированного) холестерола с липопротеиновых частиц, которые существенно отличаются друг от друга по липидному и апопротеиновому составу, размерам и функциям. Образуются ЛПВП в печени. Оттуда они секретируются в кровоток в "незрелом" виде, то есть они имеют дисковидную форму. Такая форма обусловлена отсутствием у них ядра из нейтральных липидов. Основным их липидным компонентом являются фосфолипиды
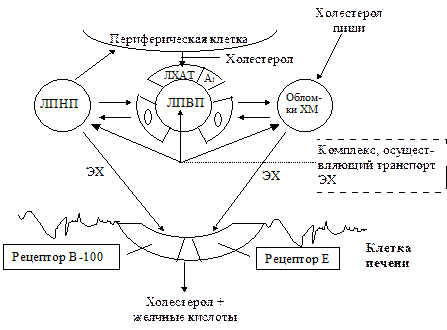
Основные пути транспорта ХС в организме
Переход свободного холестерола из клеток на ЛПВП обусловлен разницей его концентраций на поверхности клеточных мембран и липопротеиновых частиц. Следовательно, он продолжается до тех пор, пока не выровняется концентрация холестерола между донором (поверхность мембран) и акцептором (ЛПВП). Поддержание градиента концентрации обеспечивается постоянным превращением свободного холестерола, поступающего на ЛПВП, в эфиры холестерола. Эта реакция, как уже указывалось, катализируется ферментом лецитин-холестерол-ацилтрансферазой (ЛХАТ[8]).
Происхождение свободного и эстерифицированного ХС в плазме крови
|
Липопротеин |
Свободный холестерол |
Эфиры холестерола |
|
ХМ |
Синтез в слизистой кишечника |
Свободный холестерол |
|
ЛПОНП |
Гепатоциты |
АХАТ гепатоцитов |
|
ЛПНП |
Гепатоциты |
АХАТ гепатоцитов - 60% ЛХАТ плазмы - 40% |
|
ЛПВП |
Периферические клеточные мембраны - 70% ЛПОНП, ЛПНП - 30% |
ЛХАТ плазмы - 100% |
гидрофобными соединениями (в отличие от свободного холестерола, у которого имеется гидроксильная группа, сообщающая ему гидрофильность). В силу своей гидрофобности эфиры холестерола теряют способность к диффузии и не могут вернуться обратно в клетку. Они формируют гидрофобное ядро внутри частиц, благодаря которому ЛПВП приобретают сферическую форму. Здесь, в кровотоке из таких ЛПВП осуществляется транспорт вновь образованных неполярных ЭХ из ЛПВП на ХМ, ЛПОНП, ЛППП, который ускоряется за счет белков-переносчиков (см. выше). Затем происходит опосредованное рецепторами поступление ЛПВП, как и других богатых ЭХ липопротеинов, в гепатоциты с последующим их катаболизмом. В частности, высвобождающиеся эфиры холестерола служат исходным субстратом для образования желчных кислот. Аккумуляция холестерола в сосудистой стенке происходит вследствие дисбаланса между поступлением его в интиму сосудов и его выходом. В результате такого дисбаланса холестерол там накапливается. В центрах накопления холестерола формируются структуры - атеромы. Наиболее известны два фактора, которые вызывают дисбаланс в обмене холестерола. Во-первых, это изменения частиц ЛПНП (гликозилирование, перекисное окисление липидов, гидролиз фосфолипидов, окисление апо В). Поэтому они захватываются специальными клетками - "мусорщиками" (главным образом, макрофагами). Захват липопротеиновых частиц с помощью "мусорных" рецепторов протекает бесконтрольно. В отличие от апо В/Е - опосредованного эндоцитоза это не вызывает регуляторных эффектов, направленных на снижение поступления в клетку ХС, описанных выше. В результате макрофаги переполняются липидами, теряют функцию поглощения отходов и превращаются в пенистые клетки. Последние задерживаются в стенке кровеносных сосудов и начинают секретировать факторы роста, ускоряющие клеточное деление. Возникает атеросклеротическая пролиферация клеток.
Организационно- методические указания лаборантскому составу:
Подготовить мультимедийную презентацию лекции в Power Point и flash проекциях
Лекция подготовлена доц. каф. Свергун В.Т.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.