Каждая хромосома имеет первичную перетяжку, разделяющую хромосому на два плеча. В зоне первичной перетяжки располагается центромера или кинетохор. Правда, некоторые авторы придают в настоящее время термину кинетохор другое значение: так называют появляющуюся в мета- и анафазе в области центромеры зону, ответственную за взаимодействие с «кинетохорными микротрубочками» ахроматинового веретена.
Кроме первичной различают еще и вторичную перетяжку, которая чаще всего отделяет небольшой кусочек плеча хромосомы, называемый спутником. Такие хромосомы называют спутничными, а зона вторичной перетяжки зачастую содержит ядрышковый организатор и является местом формирования ядрышка.
По местоположению центромера в цитологии хромосомы делятся на следующие типы:
1) метацентрические – оба плеча равны;
2) субметацентрические – одно плечо больше другого;
3) акроцентрические – одно плечо очень маленькое;
4) телоцентрические – второе плечо практически неразличимо.
Таким образом, с учетом общей длины хромосом и их плеч, местоположения первичной перетяжки и центромеры, а также наличия вторичной перетяжки морфология большинства хромосом различается. Это и служит основой для изучения кариотипа для каждого вида и составления кариограммы. Однако зачастую этих параметров бывает недостаточно, особенно когда возникает необходимость легко и быстро идентифицировать отдельные хромосомы. По общей морфологии это сделать очень трудно, поэтому проводят дифференциальную окраску хромосом – специфическое окрашивание определенных зон хромосом (например, окрашивание для выявления А-Т-богатых зон хромосом или наоборот зон богатых Г-Ц-парами азотистых оснований). В настоящее время существует достаточно широкий спектр различных методов окрашивания, которые позволяют выявлять только специфические для каждого метода участки хромосом. Возникает полосатая исчерченность хромосом – характерная картина дифференциального окрашивания хромосом.
Политенные хромосомы слюнных желез дрозофилы (и других двукрылых насекомых) – это идеальная модель для изучения как морфологии, так и функционирования хромосом. Под обычным микроскопом они видны в интерфазном ядре в гаплоидном количестве (ибо находятся в конъюгированном состоянии, как в мейозе) в виде крупных тяжей, состоящих из чередующихся темных и светлых поперечных полос (дисков) с расширениями (пуфами) в некоторых участках.
Само название политенных (поли – много, тенос – нить) хромосом связано с наличием в их составе вместо двух хроматид 1024 и более (что является результатом 9–10 циклов репликаций хромосомы). Именно поэтому, в отличие от всех прочих интерфазных хромосом, они и видны в обычном микроскопе в неделящемся ядре и их можно даже картировать, можно различать активно работающие зоны – пуфы, и зоны блокированные – гетерохроматиновые. Всего различают у дрозофилы около 5000 темных дисков, разделенных светлыми междисковыми районами. Местоположение и морфология их стабильны, что позволяет составить цитологическую карту хромосомы. С учетом того, что многие диски соответствуют генам или блокам генов, можно с некоторой степенью уверенности говорить об интерфазном картировании генов. При специфическом активировании некоторых генов в строго определенных участках происходит формирование пуфов – деспирализация ДНК в связи с ее транскрипцией, т. е. происходит визуализация гена в процессе его работы.
Синтез ДНК называется репликацией или редупликацией (удвоением), синтез РНК – транскрипцией (переписывание с ДНК). Синтез белка, проводимый рибосомой на матричной РНК, называется трансляцией, то есть, переводим с языка нуклеотидов на язык аминокислот.
Молекула ДНК (рис. 1.7), состоящая из двух спиралей, удваивается при делении клетки. Удвоение ДНК основано на том, что при расплетении нитей к каждой нити можно достроить комплементарную копию, таким образом получая две нити молекулы ДНК, копирующие исходную.
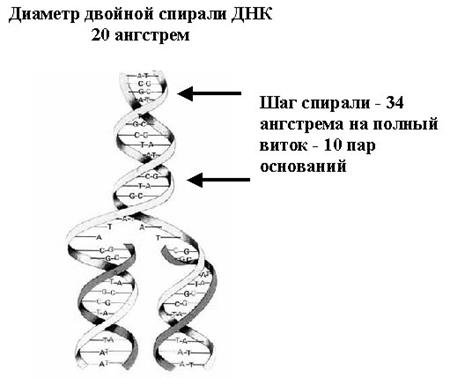
Рис. 2.10. Молекула ДНК
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.