





















Лимфопоэз у эмбриона и плода происходит транзиторно в различных эмбриональных тканях. Лимфоидные предшественники присутствуют уже в желточном мешке. К 5-6 неделе гестации предшественники B- и T-клеток появляются в печени, где формируются участки B-лимфопоэза. Другие эмбриональные ткани, включая сальник и плаценту, также содержат предшественники В-клеток.
Селезенка становится очагом гемопоэза в третьем триместре беременности и остается местом гемопоэза до момента рождения. У взрослых селезенка функционирует как вторичный лимфоидный орган, в котором располагаются зрелые T- и B-клетки. После рождения костный мозг становится основным местом развития миелоидных клеток и B-лимфоцитов.
Формирование большинства T-клеток происходит в тимусе. На 7-8 неделе эмбрионального развития формирующийся тимус заселяется принесенными с кровью предшественниками, у взрослых в этот орган перемещаются костномозговые предшественники, где из них развиваются Т-лимфоциты, а процесс называется Т-лимфопоэзом.
В настоящее время предполагается, что предшественники, коммитированные к B- и T-клеточному пути развития, являются прямыми потомками плюрипотентной стволовой клетки (CD34+).
Развитие лимфоцита у взрослого проходит две фазы. Первая фаза – антигеннезависимая, когда B-лимфоциты развиваются до стадии, в которой они экспрессируют на поверхности иммуноглобулин (Ig) М, происходит в костном мозге. Вторая фаза – антигензависимая – протекает во вторичных лимфоидных органах, таких как селезенка и лимфатические узлы, в ответ на клеточные и гуморальные сигналы, поступающие от T-клеток, макрофагов и других популяций акцессорных клеток.
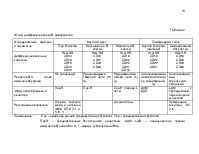
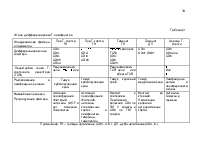
При использовании специфических моноклональных антител, которые распознают иммуноглобулины и другие поверхностные и цитоплазматические детерминанты, можно различать промежуточные степени развития B-клеток (таблица 1).
Клетки-предшественники, коммитированные к дифференцировке в B-лимфоциты, имеют иммуноглобулиновые гены в зародышевой конфигурации и называются про-B-клетки. По мере созревания этих клеток они экспрессируют на поверхности антиген CD45RA, тирозинфосфатазу, чья функция в развитии B-клетки пока неизвестна. Затем в цитоплазме появляется белок тяжелой цепи иммуноглобулина М (), и с этого момента клетка обозначается как пре-B-клетка. Как только в клетке происходит экспрессия белков легкой цепи, начинается сборка молекул иммуноглобулинов, и после их появления на поверхности клетки определяются как B-лимфоциты.
Наиболее незрелая человеческая B-клетка-предшественник экспрессирует CD34 и CD19. Последний антиген представлен на B-клетках всех стадий развития. На стадии клетки-предшественника происходит потеря экспрессии CD34, и на поверхности клеток появляется антиген CD10. Далее пре-B-клетки, определяемые по присутствию цитоплазматического протеина , экспрессируют CD20. В течение перехода от пре-B к B-клетке происходит потеря CD10 и начинается сборка CD21, CD22 и поверхностного иммуноглобулина М (рис.1).
B-лимфопоэз характеризуется высокой клеточной пролиферацией, которая наиболее активна на стадии клеток-предшественников. К тому времени, когда клетки созревают до малых пре-B-лимфоцитов, которые экспрессируют цитоплазматический протеин , деление клеток прекращается. Однако имеется и значительная потеря клеток на стадии пре-B-лимфоцитов. Удаляются клетки, в которых произошла дефектная реаранжировка (перестройка) иммуноглобулиновых генов или экспрессируются аутореактивные гены тяжелых цепей. Клетки, которые прогрессируют до стадии экспрессии IgM, покидают костный мозг и мигрируют в селезенку, где они подвергаются дальнейшему созреванию. Если эти клетки не получают антигенного стимула, они проживают только несколько дней, после чего подвергаются апоптозу.
Предполагается, что существуют три различные линии B-клеток, определяемые по фенотипическим и функциональным свойствам: B1a, B1b и В2. B1a-клетки отличаются том, что экспрессируют CD5, B1b-клетки не экспрессируют CD5, а характеризуются высокими уровнями поверхностного IgM и низкими уровнями мембранного IgD. Распространение этих популяций у взрослых в значительной степени ограничено перитонеальной и плевральной полостями, причем В1-клетки секретируют преимущественно антитела IgA и IgM, которые обеспечивают защиту против окружающей микрофлоры. Этим они отличаются от стандартных (или B2) B-клеток, которые преобладают во вторичных лимфоидных органах, таких как лимфоузлы и селезенка.
Характерной особенностью B1-клеток является их способность производить аутоантитела класса IgM, что делает понимание их природы клинически важным вопросом. Популяция B1-клеток является, вероятно, основным источником естественных аутоантител, представленных в сыворотке крови здоровых лиц. Естественные аутоантитела встречаются в крови взрослых, детей, новорожденных и даже у плода. Они присутствуют в малых количествах и почти всегда связываются со многими антигенам (мультиреактивные аутоантитела) и имеют низкую аффинность к их лигандам. Пока не определено однозначно, продуцируются ли естественные аутоантитела исключительно B1-клетками, или их способны производить также B2-клетки.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.