|
Некоторые факторы трансляции эукариот |
|
|
Факторы эукариот |
Функции |
|
Инициация |
|
|
еIF1 |
Участвуют в образовании инициирующего комплекса |
|
eIF2 |
|
|
eIF3, eIF4C |
|
|
eIF4E |
Участвует в связывании кэпа иРНК |
|
eIF4A, eIF4B,eIF4F |
Участвуют в поиске первого кодона АУГ |
|
eIF5 |
Способствуют диссоциации eIF2, eIF3, eIF4C |
|
eIF6 |
Способствует диссоциации 60S субъединиц |
|
Элонгация |
|
|
eEF1a |
Доставляет аминоацил тРНК к рибосоме |
|
eEF1bg |
Участвует в рециклизации eEF1a |
|
eEF2 |
Фактор транслокации |
|
Терминация |
|
|
eRF |
Завершает синтез полипептидной цепи |
Наибольшим сродством к малой субъединице рибосомы обладает IF3 , котрорый является ведущим белком, поддерживающим эту субъединицу в свободном нативном состоянии. Другой фактор, IF2, является представителем большой группы белков, способных связываться с гуаниловыми нуклеотидами и обладающими ГТФазной активностью. Он вступает в процесс на следующем этапе инициации, который может осуществляться двумя способами
а) нативная малая субъединица вначале взаимодействует с иРНК, и затем к этому комплексу присоединяется инициирующая аминоацил-тРНК связанная с IF2 и ГТФ
б) с малой субъединицей вначале взаимодействует инициирующая аминоацил-тРНК с IF2 и ГТФ, а затем к этому комплексу присоединяется иРНК
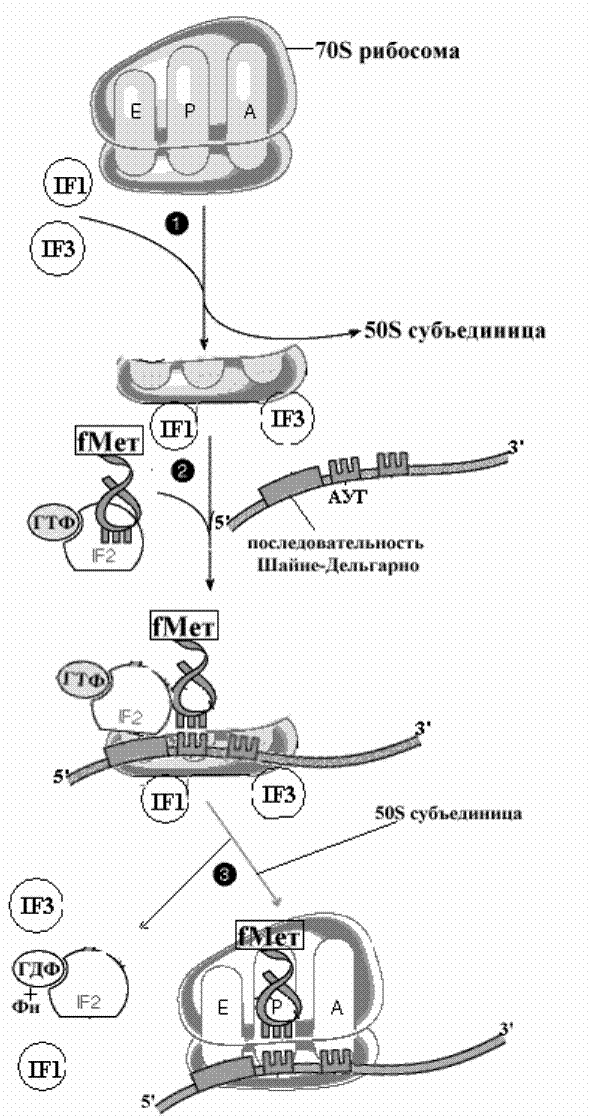 |
Рис..Образование инициирующего комплекса у прокариот.
Как видно и тот и другой способы приводят к образованию так называемого инициирующего комплекса, состоящего из малой рибосомной субъединицы, иРНК, инициирующей аминоацил-тРНК и факторов инициации. Образование инициирующего комплекса решает важную задачу точной установки инициирующей аминоацил тРНК в начале кодирующей последовательности иРНК.
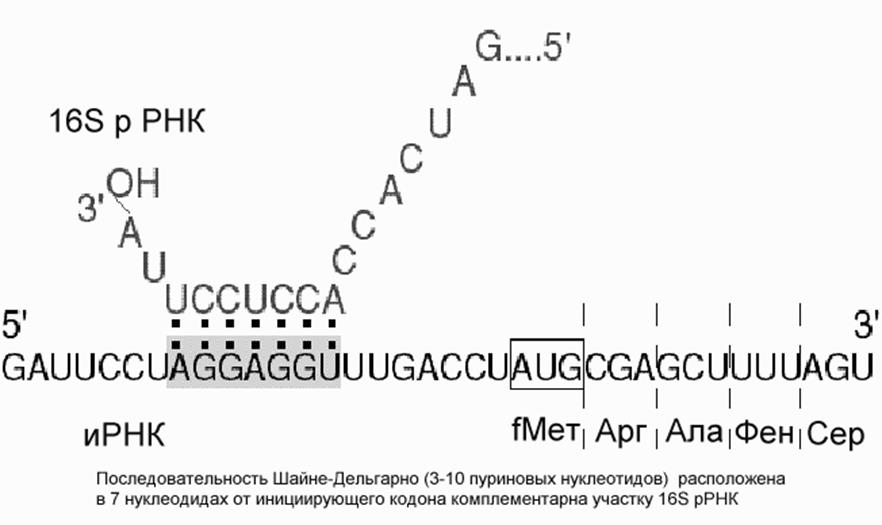 |
У прокариот и у эукариот это происходит по-разному. У прокариот малая рибосомная субъединица с факторами инициации имеет повышенное сродство к специальному внутреннему участку иРНК, который называют инициирующим, или рибосомосвязывающим участком.. Наиболее известной универсальной структурой такого участка является полипуриновая нуклеотидная последовательность Шайне-Дельгарно , отстоящая от инициирующего кодона на несколько (3-10) нуклеотидов к 5'-концу, т.е. предшествующей инициирующему кодону.
Эта последовательность в большей или меньшей степени комплементарна 3'-концевому участку рибосомной 16S РНК, богатому пиримидиновыми нуклеотидными остатками (5'-GAUCACCUCCUUA-3' у E.coli). Комплементарными могут быть от 3 до 9 остатков (обычно около 4-5). Последовательность CCUCC 3'-концевого участка 16S РНК E.coli оказывается наиболее часто комплементарной (хотя, как правило, частично) полипуриновой прединициирующей последовательности иРНК. Считается, что комплементарное спаривание 3'-концевого участка 16S РНК с прединициирующим полипуриновым блоком иРНК и обеспечивает точную установку инициирующего комплекса на начало кодирующей последовательности иРНК.
У эукариот малая рибосомная субъединица с факторами инициации узнает кэп 5'-концевого участка иРНК, и как правило, взаимодействует с ним. У эукариот малая субъединица обычно взаимодействует с инициирующей аминоацил-тРНК до связывания иРНК. Связавшись с 5'-концом иРНК вместе с инициирующей аминоацил-тРНК и факторами инициации, малая рибосомная субъединица начинает двигаться от 5'-конца по направлению к 3'-концу, используя энергию гидролиза АТФ для раскручивания вторичной структуры иРНК. АТФазной и хеликазной активностью обладает специальный эукариотический фактор инициации eIF4. Когда рибосомная частица достигает и узнает инициирующий кодон, антикодон инициирующей аминоацил-тРНК взаимодействует с ним, обеспечивая тем самым узнавание начала кодирующей последовательности иРНК .
 Единственным инициирующим кодоном эукариот является триплет АУГ. У
прокариот с этого триплета также начинается 90% всех кодирующих
последовательностей. Однако некоторая часть прокариотических мРНК инициирует
трансляцию на других триплетах - ГУГ, реже УУГ и совсем редко на некоторых
других . Очевидно, что у прокариот способность некоторых триплетов быть
инициирующими определяется их положением в инициирующем или
рибосомосвязывающем участке иРНК . Эти же триплеты, когда они встречаются в
кодирующей последовательности в ходе элонгации, кодируют ту или иную аминокислоту
АУГ - метионин, ГУГ - валин, УУГ - лейцин .
Единственным инициирующим кодоном эукариот является триплет АУГ. У
прокариот с этого триплета также начинается 90% всех кодирующих
последовательностей. Однако некоторая часть прокариотических мРНК инициирует
трансляцию на других триплетах - ГУГ, реже УУГ и совсем редко на некоторых
других . Очевидно, что у прокариот способность некоторых триплетов быть
инициирующими определяется их положением в инициирующем или
рибосомосвязывающем участке иРНК . Эти же триплеты, когда они встречаются в
кодирующей последовательности в ходе элонгации, кодируют ту или иную аминокислоту
АУГ - метионин, ГУГ - валин, УУГ - лейцин .
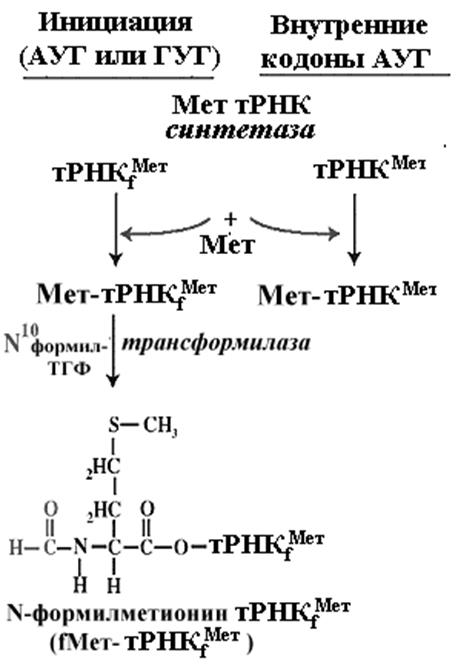
И у прокариот и у эукариот инициирующей аминоацил-тРНК является метионил-тРНК, у прокариот, как уже отмечалось, ее аминогруппа блокирована формильным остатком -формил-метионил-тРНК. Ее антикодон всегда ЦАУ, так что он полностью комплементарен кодону АУГ и частично комплементарен другим возможным инициаторным кодонам прокариот [например, ГУГ и УУГ].
Структура молекулы тРНК выпонящей роль инициирующей метионил-тРНК несколько отличается от структур других тРНК, в том числе и от структуры метионил-тРНК, используемой в процессе элнгации, что и делает ее функцию в качестве инициатора трансляции уникальной. Инициаторные тРНК имеют консервативный участок ГАУЦ в положении 51-54. Универсальная последовательность в тех же позициях элонгаторных тРНК содержит T вместо A и псевдоуридин вместо У. В инициаторных тРНК дигидроуридиловая петля не содержит дигидроуридина Особенности структуры инициирующей метионил-тРНК обеспечивают ее специфическое комплексирование с белком IF2 при наличии ГТФ [ГТФ, присоединяясь к белку, повышает сродство белка к инициирующей метионил-тРНК] . Универсальность инициации с участием инициирующей метионил-тРНК приводит к тому, что всегда и
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.