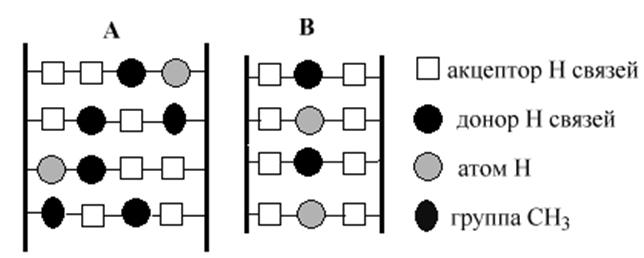
Модель пространственной структуры ДНК предложенная Криком и Уотсоном в 1953 г на первый взгляд казалась монотонно устроенной структурой с жестко детерминированными расстояниями и углами поворота между основаниями. Однако более детальное исследование показало существование локальных особенностей структуры, позволяющих специфически узнавать эти особенности молекулам белков. Химические группы пар азотистых оснований хорошо «просматриваются» в большой и малой бороздках молекулы ДНК, создавая неповторимые сочетания, которые могут выполнять роль специфических знаков для узнавания белками.
 |
|
. Белки регуляторы и узнаваемая ими последовательность нуклеотидов на ДНК |
||
|
Белок |
Последовательность |
|
|
E.Coli |
Lac-репрессор |
ААТТГТГАГЦГГАТААЦААТТ ТТААЦАЦТЦГЦЦТАТТГТТАА |
|
САР |
ТГТГАГТТАГЦТЦАЦТ АЦАЦТЦААТЦГАГТГА |
|
|
Дрожжи |
GAL-4 |
ЦГГАГГАЦТГТЦЦТЦЦГ ГЦЦТЦЦТГАЦАГГАГГЦ |
|
Животные |
Oct-1 |
АТГЦАААТ ТАЦГТТТА |
|
GATA-1 |
ТГАТАГ АЦТАТЦ |
|
В основе молекулярного узнавания лежит принцип комплемнтарности между поверхностями двух молекул. В случае взаимодействия регуляторного белка и молекулы ДНК между поверхностью белка и структурой ДНК образуется множество специфических контактов (по меньшей мере, в 20 точках) при участии водородных связей, ионного и гидрофобного взаимодействий. Слабые взаимодествия в таком количестве делают это взаимодействие достаточно прочным. Исследование структуры многих белков, участвующих в регуляции экспрессии генов, позволило выделить несколько групп белков, объединенных общими признаками в структурной организации.
Спираль-поворот-спираль. Первой изученной связывающей ДНК структурой в белке была структура названная спираль- поворот- спираль. Подобная структура, обеспечивающая специфическое связывание белка и ДНК обнаружена в сотнях ДНК связыващих белков и у прокариот и у эукриот. 2 спиральных участка молекулы согнутые под углом в области короткого линейного отрезка располагались таким образом, что С-концевой отрезок вписывался в большую бороздку ДНК и специфически узнавал определенную последовательность нуклеотидов. Как правило, все белки с такой структурой выполняли свою функцию в виде димеров. К этой группе белков относятся и гомеодомен: гомеодомен - высоко консервативный домен из 60 аминокислот, обнаруженный среди большого семейства факторов транскрипции. Это семейство было сначала идентифицировано у Drosophila (до 60 белков), а впоследующем обнаружены у всех живых организмов от дрожжей до человека. Эти белки кодируются группой генов, изменение которых, может вызвать нарушение номального формирования организма ( преобразования одной части тела в другую, так называемые гомеотические преобразования). Этот класс генов был идентифицирован как у беспозвоночных так и позвоночных. Сам гомеодомен формирует структуру, высоко подобную бактериальным белкам типа спираль-поворот-спираль.
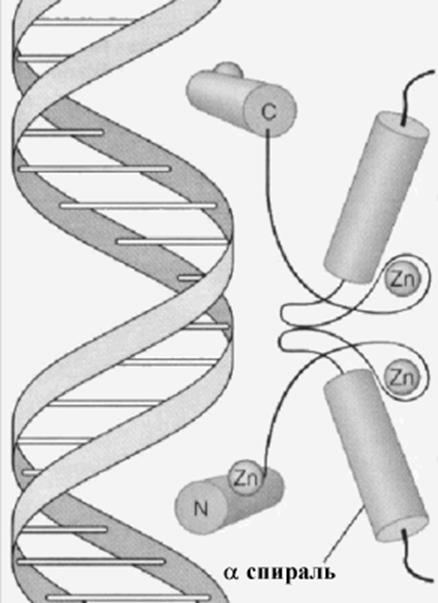 Цинковые пальцы: Если структура спираль-поворот-спираль
состоит только из аминокислот, второй тип наиболее часто встречающихся типов
структур узнающих ДНК содержит в качестве стабилизирующего элемента ион цинка.
Существует целое семейство таких белков, наиболее простой из которых составлен
из a спирального отрезка
цепи и отрезка с b -
структурой, связанных между собой ионами цинка. Этот тип белков участвует в
регуляции транскрипции рибосомных РНК (фактор транскрипции РНК pol III, TFIIIA). Второй вариант
такой структуры составлен из 2-х a-
спиральных участков, также связанных ионами цинка. К ним относятся внутриклеточные
рецепторы стероидных гормонов и гомонов щитовидной железы. Атомы цинка
связываются с остатками цистеина и гистидина, способствуя формированию пальцевидных
структур. И тот и другой варианты используют a -спиральные структуры для узнавания участков. Области пальца
могут "вставляться " в главное углубление спирали ДНК. Интервал
цинковой области пальца в этом классе факторов транскрипции совпадает с полуповоротом
двойной спирали.
Цинковые пальцы: Если структура спираль-поворот-спираль
состоит только из аминокислот, второй тип наиболее часто встречающихся типов
структур узнающих ДНК содержит в качестве стабилизирующего элемента ион цинка.
Существует целое семейство таких белков, наиболее простой из которых составлен
из a спирального отрезка
цепи и отрезка с b -
структурой, связанных между собой ионами цинка. Этот тип белков участвует в
регуляции транскрипции рибосомных РНК (фактор транскрипции РНК pol III, TFIIIA). Второй вариант
такой структуры составлен из 2-х a-
спиральных участков, также связанных ионами цинка. К ним относятся внутриклеточные
рецепторы стероидных гормонов и гомонов щитовидной железы. Атомы цинка
связываются с остатками цистеина и гистидина, способствуя формированию пальцевидных
структур. И тот и другой варианты используют a -спиральные структуры для узнавания участков. Области пальца
могут "вставляться " в главное углубление спирали ДНК. Интервал
цинковой области пальца в этом классе факторов транскрипции совпадает с полуповоротом
двойной спирали.
Спираль-петля-спираль (HLH): HLH домен формирует димеры. HLH домен составлен из двух a- спиральных областей , разделенных областью переменной длины, которая формирует петлю между 2 a – спиралями. Этот домен подобен структуре спираль-поворот-спираль, a спиральные области структурно подобны и необходимы для взаимодействия белка с элементами последовательности, которые обладают двойной осью симметрии. Этот класс факторов транскрипции наиболее часто содержит область основных аминокислот, расположенных на N – концевом участке HLH домена (такие белки обозначают как bHLH белки). Эти аминокислоты необходимы для связывания с ДНК по специфическим последовательностям. HLH область функционирует в форме гомо и гетеродимеров. Среди белков, имеющих HLH домен можно назвать MyoD ( фактор транскрипции идуцирующий миогенез) и c-Myc (первоначально идентифицированный как ретровирусный онкоген). Некоторые HLH белки не содержат основной области и, образуя гетеродимеры с белками bHLH, действуют как репрессоры, подавляя активность bHLH белков.
Лейциновая застежка - молния: как указывалось выше большинство белков, взаимодействуют с ДНК в форме димеров, причем в образовании димера участвуют последовательности аминокислот, которые не задействованы в самом механизме взаимодействия с ДНК.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.