Белки с лейциновым доменом застежки - молнии сочетают при этом обе функции и образование димера и связывание с ДНК. В белке имеются участки богатые гидрофобными аминокислотами (часто лейцином) радикалы которого выступают из a спиральной структуры этого белка. Два белка с такими участками могут формировать суперспираль в которой выступающие группы радикалов лейцина взаимодействуют с лейциновыми радикалами другого мономера подобно застежке - молнии. Димер при этом приобретает Y- форму. Ножки этой структуры содержат участки узнавания последовательности ДНК. Область лейциновой застежки - молнии присутствует в многих связывающих ДНК белках, типа c-Myc, C/EBP и т.д.
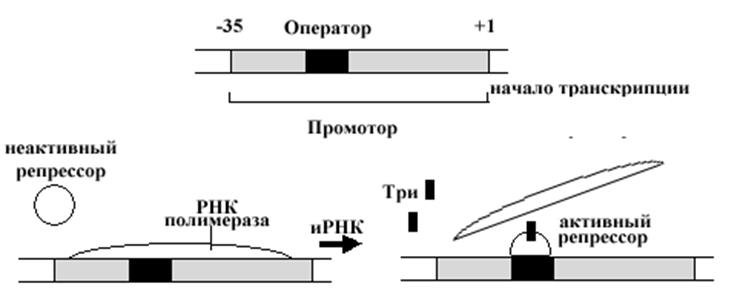 |
Рис..Схема влияния триптофана на работу оперона.
ваются с соответствующими операторными участками и блокируют продвижение РНК-полимеразы, и, следовательно, препятствуют транскрипции. В самом простом варианте этот механизм можно рассмотреть на примере триптофанового оперона кишечной палочки. Оператор триптофанового оперона представляет собой последовательность нуклеотидов, узнаваемую репрессором этого оперона, относящегося к белкам со структурой спираль-петля-спираль. Присоединение репрессора структурно блокирует доступ к промотору РНК-полимеразы, предотвращая экспрессию триптофанового оперона. Присоединение репрессора к оператору становится возможным лишь в том случае, если к репрессору присоединятся 2 молекулы триптофана.
Молекулы триптофана изменяют конформацию репрессора, обеспечивая возможность его a -спиральным участкам соединиться с группами нуклеотидов большой бороздки ДНК в области оператора. Такое влияние продукта гена регулятора получило название негативного контроля, гены, кодирующие такие регуляторы, названы генами репрессоров, а молекулы, способствующие такой реакции репрессора, получили название корепрессоры. Роль корепрессоров в клетке часто выполняют конечные продукты метаболических путей.
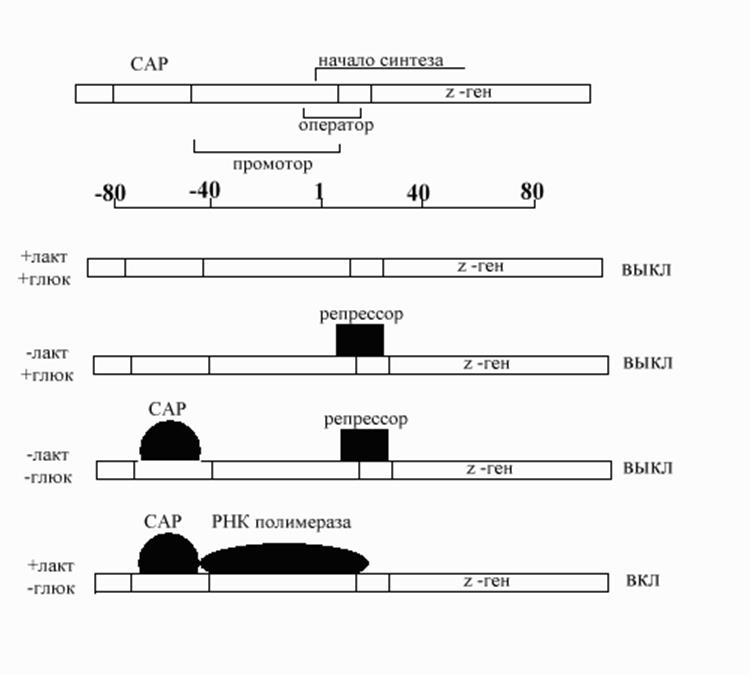 |
кишечной палочки. lac оперон состоит из одного гена-регулятора (i ген) и трех структурных генов (z, y, и a). Ген i кодирует репрессор lac оперона. Z ген кодирует b-галактозидазу, ответственную за гидролиз дисахарида лактозы на ее мономерные единицы, галактозу и глюкозу. Y ген кодирует пермеазу, которая увеличивает проницаемость клетки к b-галактозидам, а ген a - трансацетилазу. При выращивании кишечной палочки на среде, содержащей глицерол в качестве единственного источника углерода, lac репрессор связан с оператором и РНК-полимераза не может присоединиться к оператору. Оперон выключен. Лактоза, добавленная в среду, связывается с репрессором и резко снижает его способность связываться с оператором. Это обеспечивает присоединение РНК-полимеразы к промотору, но синтез РНК становится возможным лишь только в том случае, когда к участку ДНК, расположенному выше (к 5’концу) промотора присоединится еще один белок, названый САР (катаболитами активируемый белок). Этот белок является рецептором цАМФ, уровень которой в клетке определяется уровнем глюкозы (снижение количества глюкозы в питательной среде приводит к повышению уровня цАМФ в клетке). САР, связанный с цАМФ, присоединяется к ДНК и стимулирует РНК-полимеразу, при этом ее активность увеличивается в 20 –50 раз.
Все возможные варианты ответа оперона на изменения компонентов питательной среды показаны на рис.10-21. В отличие от триптофанового оперона, где роль включателя оперона выполнял один сигнал (триптофан), в случае lac оперона требуются 2 сигнала, причем лактоза оказывала положительный эффект, а глюкоза – пример негативного контроля. Небольшие молекулы, которые подобно лактозе могут связываться с белками-репрессорами и ингибировать их способность связываться с операторными участками ДНК получили название индукторы. Индукторами транскрипции служат субстраты метаболических путей. Они стимулируют синтез белков, обеспечивающих их превращения.
Значительно более сложными являются механизмы регуляции инициации транскрипции у эукариот. Можно выделить 2 важнейшие стороны такого различия. Во-первых, РНК-полимераза эукариот не может сама катализировать синтез РНК без участия специальных белков –факторов транскрипции, которые должны присоединиться к промотору и самой полимеразе, при этом существует довольно широкие возможности по управлению скоростью транскрипции путем сочетания этих факторов. Вторая отличительная особенность регуляции инициации транскрипции у эукариот это то, что факторы регуляции могут оказывать влияние на промотор, присоединяясь к ДНК на расстояниях в тысячи пар оснований от промотора, что позволяет оказывать влияние на один промотор множеству регуляторных белков. далеких расстояниях от промотора,
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.