Как уже отмечалось в разделе о механизмах синтеза РНК существуют довольно значительные различия в структуре регуляторной части участка ДНК, которая используется для транскрипции у эукариот и прокариот. На рисунке показаны некоторые факторы транскрипции и возможный порядок их присоединения, необходимый для проявления активности РНК-полимеразы II. Присоединению РНК-полимеразы к промотору, который представлен последовательностью нуклеотидов, богатых Т и А (ТАТА –бокс), предшествует присоединение
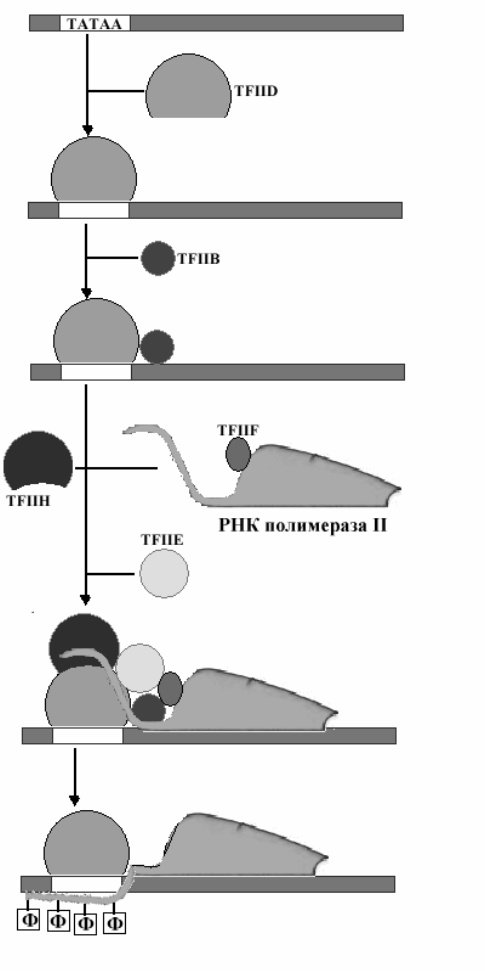 |
специального белкового фактора TFIID. Этот фактор состоит из нескольких субъединиц, одна из которых специфически связывается с ТАТАА последовательностью (ТАТА связывающий белок- TBP).
Вслед за TFIID к промотору начинают присоединяться другие белки и РНК- полимераза. Один из белковых факторов(TFIIH) обладает протеинкиназной активностью и после завершения образования комплекса катализирует фосфорилирование РНК-полимеразы, что и является сигналом для включения транскрипции.
Процессы формирования комплекса факторов инициации регулируются специальными регуляторными белками, которые присоединяются к специфическим последовательностям, как правило, расположенным на больших расстояниях от промотора. В свою очередь, регуляторные белки, связывающиеся с ДНК формируют сложные ансамбли, конечный эф
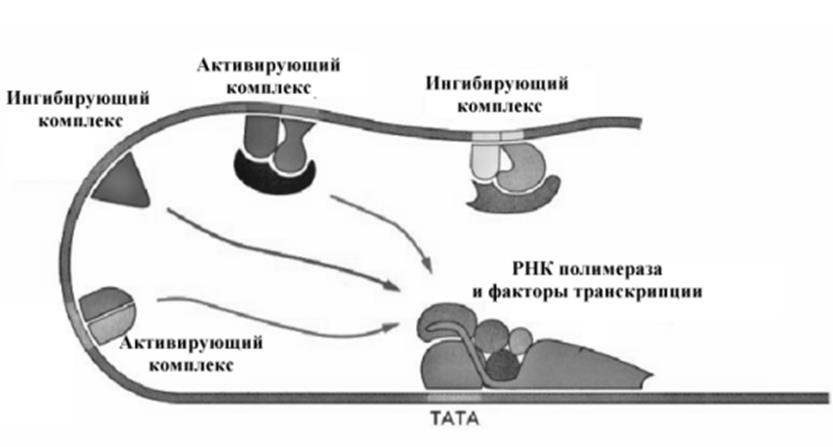 |
фект действия которых зависит от соотношения белков в ансамбле. Влияние на процесс формирования комплекса инициирующих белков может быть ускоряющим (энхансеры) или замедляющим ( сайленсеры). Регуляторные белки имеют как минимум два специфических домена, один из которых обеспечивает узнавание и взаимодействие с ДНК, а второй домен обеспечивает взаимодействие с другими молекулами (белками, метаболитами, гормонами и т.д.). Строение домена, обеспечивающего взаимодействие с ДНК описано выше. В свою очередь конформация самих факторов регуляции регулируется целым рядом механизмов, некоторые из них представлены на рис.10-24.
Важным элементом в механизмах регуляции экспрессии генов на уровне транскрипции является доступность участков ДНК к действию регуляторов транскрипции. Это связано прежде всего со способом упаковки генетического материала в форме комплекса с белками –хроматина, что предполагает необходимость деконденсации хроматина как первого шага на пути регуляции транскрипции. Многие гены, используемые для синтеза белков собраны в
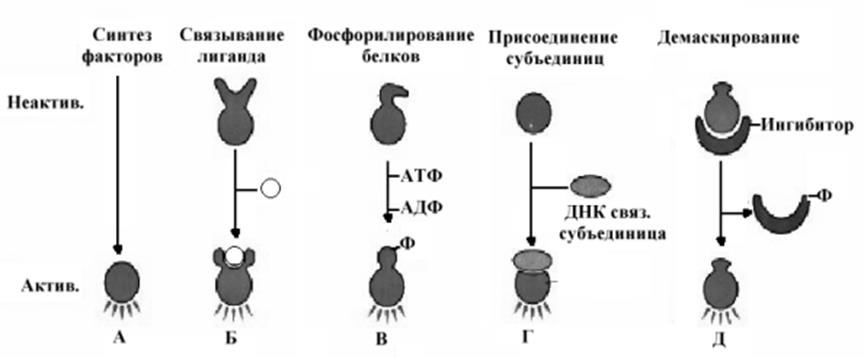 |
молекулах ДНК в форме кластеров и доступность к такому кластеру может быть в свою очередь регулируема. Предполагается несколько механизмов, позволяющих вызвать изменения хроматина. Возможно, существуют белки, катализирующие изменения хроматина, или являющиеся ингибиторами деконденсации и их удаление способствует этому процессу. Включение или выключение отдельных кластеров генов в определенные сроки жизни клетки или организма обеспечивают процессы дифференцировки клеток, адаптации к определенным условиям жизни и т.д.
Основным объектом регулирующего влияния на синтез белка и нуклеиновых кислот является транскрипция. Однако существует возможность регуляции и после транскрипции. Это происходит на всех этапах процессинга РНК, трансляции и посттрансляционной модификации белковых молекул.
У прокариот существует возможность остановить начавшуюся транскрипцию, благодаря специальной последовательности нуклеотидов на РНК, которая обеспечивает удаление РНК-полимеразы с матрицы ДНК. Это хорошо показано на примере упоминаемого выше триптофанового оперона кишечной палочки. У оперона триптофана имеется вторая регуляторная область, область аттенуации (ослабления).
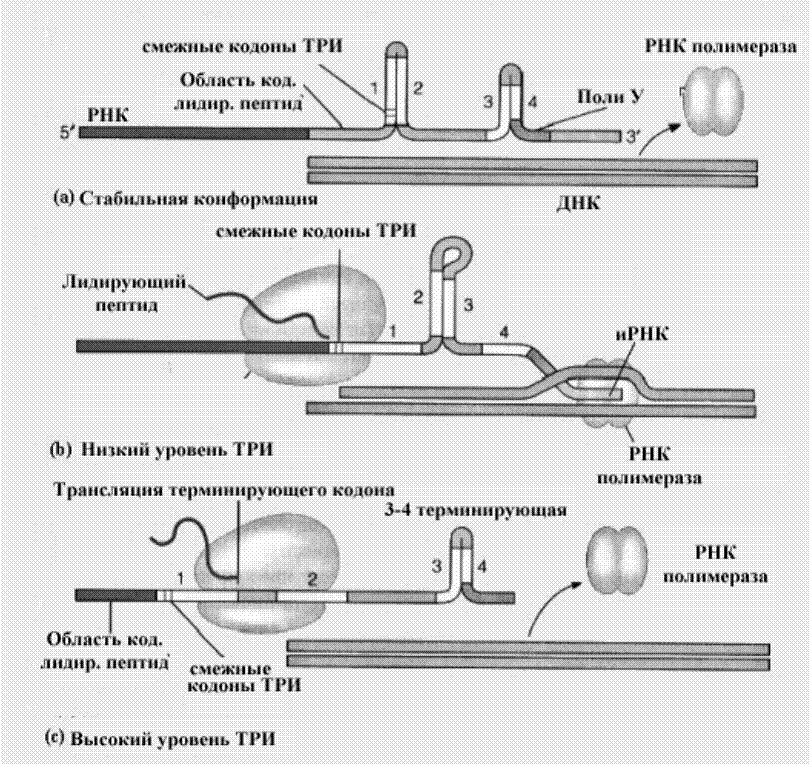
Этот участок, расположен приблизительно на 140 нуклеотидов вниз
к 3’концу от участка инициации иРНК. Синтез иРНК может быть преждевременно
остановлен на этом участке, если уровень триптофана повышается выше некоторого
значения. Механизм ослабления показан на рис . Лидирующий транскрипт кодирует
14-амино-кислотный олигопептид, который содержит два смежных остатка
триптофана. Он также содержит четыре облас
Рис.Аттенуация транскрипции на примере триптофанового оперона Е.coli.
ти (1, 2, 3, и 4) способные к формированию трех шпилек, SL-1:2, SL-2:3, и SL-3:4. SL-3:4 - участок терминации транскрипции; он непосредственно следует за отрезком поли У. Шпилька SL-3:4 может образоваться, если предотвратить образование шпильки SL-2:3.
В отсутствии триптофана (то есть, когда необходима полная экспрессия оперона), рибосома задерживает два кодона триптофана, расположенные в области 1. В результате, SL-1:2 не может формироваться, а вместо этого формируется шпилька SL-2:3, что в свою очередь предотвращает формирование шпильки SL-3:4, и синтез иРНК не прерывается, так как SL-2:3 не узнается как сигнал терминации транскрипции.
В присутствии триптофана, однако, рибосома переместится в область 2 лидирующего транскрипта прежде, чем та образует пары оснований с областью 3, таким образом, не допуская формирования SL:-2:3. Это способствует образованию SL-3:4, что приводит к преждевременной терминации транскрипции. Аттенуация транскрипции происходит и в случае, когда останавливается
Следует заметить, что аттенуация транскрипции, по-видимому, ведущий способ регуляции и других оперонов, привлекаемых для биосинтеза аминокислот. Лидирующие последовательности иРНК, образующиеся при транскрипции оперонов гистидина, треонина и фенилаланина содержат по 7 или 8 кодонов для соответствующей аминокислоты. Это делает аттенуацию транскрипции удобной формой регуляции активности оперонов в зависимости от уровня аминокислот в цитоплазме.
 |
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.