В начале века эукариотные фотосинтетики были разбиты на несколько основных групп отделов (=типов) по составу светочувствительных пигментов, запасных питательных веществ и структуре хроматофоров.
Недавно обнаружено, что каждая группа фотосинтетиков ранга надтипа обладает своими характеристиками жгутикового аппарата.
Следовательно:
n выделенные по пигментам группы растений являются естественными;
n по строению жгутиков можно выявить родство надтипов.
Пример:
хромофиты и бесцветные простейшие, имеющие сходную структуру жгутика, объединяются в группу Heteroconta
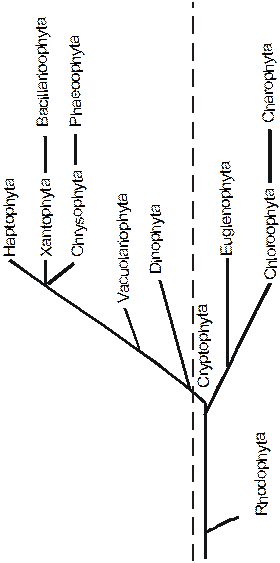
В середине века в ботанике низших растений утвердилась концепция немецкого ученого Пашера, согласно которой эукариотные фотосинтетики разбивались на 5 основных групп - Chlorophyta Cromophyta, Rodophyta, и Cryptophyta, различающиеся по набору пигментов и запасных питательных веществ (рис. 1).
Хлорофиты (зеленые водоросли) имели хлорофиллы a и b, а также набор специфических каротиноидов. Их запасными питательными веществами являлись a-4-полиглюканы (в основном представленные крахмалом). Исключение - эвгленовые, содержащие b-3-полиглюканы.
Хромофиты (золотистые, желтозеленые, бурые и диатомовые водоросли) содержали хлорофиллы а и с и другой набор каротиноидов, из которых важнейшим являлся ксантофилл фукоксантин. Исключение - динофлагелляты, имеющие специфические каротиноиды. Обе подгруппы сохраняют b-3-полиглюканы.
Родофиты (красные водоросли) несли хлорофиллы а и d, а также функционально замещавшие хлорофиллы b и с белки -, несущие особые светочувстительные пигменты фикобилины, принципиально несходные с хлорофиллами. Красные водоросли, у которых на любой стадии жизненного цикла нет никаких следов жгутикового аппарата, запасали "багрянковый крахмал" - a-4-полиглюкан, близкий к гликогену.
У криптофитов также имеются фикобилипротеины и хлорофиллы а и с. Это хищные одноклеточные организмы с двумя подвижными жгутиками и мощной клеточной глоткой, функция которой как органа захвата пищи усилена тельцами, выстреливающими липкие ядовитые нити - экструсомами.
|
Рис. 1. Схема филогении водорослей по результатам исследований Пашера и его последователей. Вертикальная прерывистая линия разграничивает группы с разными типами митохондриальных крист: слева - группы с трубчатыми кристами, справа - с пластинчатыми (Из Старобогатова, 1987) |
Ультрамикроскопические исследования 50-60-х годов показали естественность этого деления. Практически все хромофиты несут неравные жгутики. Длинный передний жгутик похож на перо - он несет два ряда трехраздельных трубчатых волосков. Короткий жгутик, часто направленный назад, голый. В основании жгутиков, в так называемой переходной зоне, имеется своеобразная спиральная структура. Их хлоропласты окружены 4 мембранами, из которых наружная оказывается частью эндоплазматической сети и часто связана с ядром. Внутри хлоропластов светочувствительные пластинки - тилакоиды - связаны в группы - ламеллы по три тилакоида в ламелле. Одна ламелла как правило огибает хлоропласт и называется поясковой ламеллой. Набор пигментов и особенности строения хлоропластов ниже будут называться "характеристиками а+с вселенца".
Динофлагелляты обладают неравными жгутиками, расположенными под углом друг к другу. Один из них - вентральный (поперечный) - имеет лентовидную форму, обусловленную причленением к типичному жгутику (микротрубочковая структура 9+2) добавочного утолщения - параксиального тяжа и односторонний ряд нетрубчатых волосков. Дорсальный (продольный) жгутик с нетрубчатыми волосками. Спиральной структуры нет. Хроматофоры обычно с характеристиками а+с вселенца.
Хлорофиты имеют как правило два одинаковых голых жгутика. В тех редких случаях, когда жгутики несут волоски, они не трубчатые и не трехраздельные. В переходной зоне находится иная, звездчатая структура. У всех жгутиконосцев жгутики закрепляются в цитоплазме своеобразной корешковой системой микротрубочек.. Корешковая система хлорофитов построена по Х+2+Х+2 типу, который отсутствует у хромофитов. Хроматофоры окружены 2 мембранами, тилакоиды собраны в ламеллы по 2-40, поясковых ламелл нет. Особенности строения и химического состава хроматофоров хлорофитов будут называться "характеристиками а+b вселенца ".
Эвгленовые исходно двужгутиковые, причем у большинства форм за пределы "клеточного рта" выходит один жгутик; если наружу выходят оба жгута, то один направлен вперед и работает в качестве плавательного, а второй жгутик направлен назад, волочащийся. В исключительных случаях (паразиты) имеются от 3 до 7 функционально равнозначных жгутиков. Жгутики покрыты нетрубчатыми волосками, которые растут однорядно или "кустиками". Собственно жгутик (микротрубочковое образование типичной структуры 9+2) по всей длине соединен с толстым цилиндром из плотно упакованных микротрубочек - параксиальным тяжем (paraxial rod). Переходная зона не имеет ни спиральной, ни звездчатой структуры. В типичном виде от базального тельца отходит один микротрубочковый корешок и еще один находится между базальными; всего три внутренних микротрубочковых корешка и один поперечноисчерченный. Хроматофоры окружены 3 мебранами, их химический состав и внутреннее устройство сходны с таковым хлорофитов. Назовем это "характеристиками вселенца хлорофитного типа".
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.