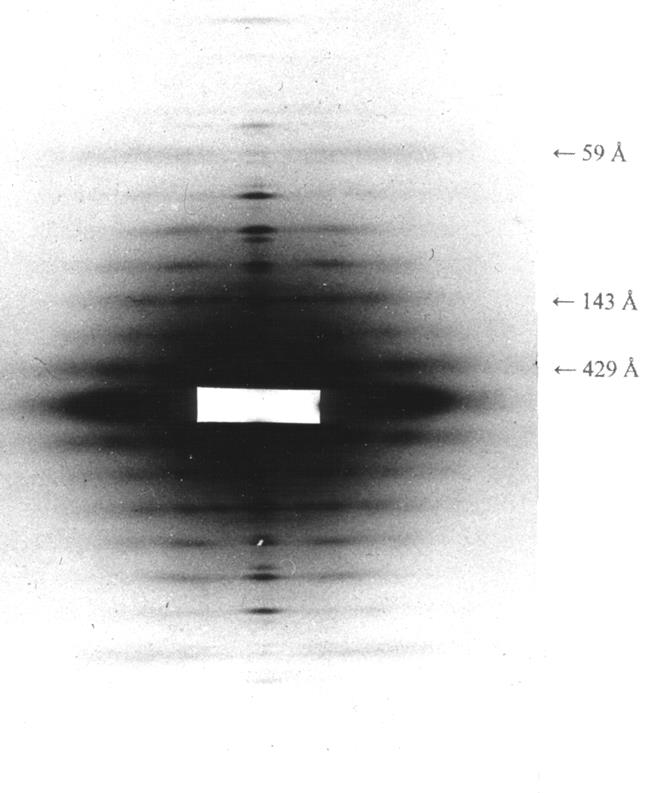
В дифракционной картине живой покоящейся скелетной мышцы, имеющей очень характерный вид в области малых углов (то есть углов, соответствующих периодам решетки, значительно большим длины волны), обычно выделяют три группы рефлексов: слоевые, меридиан и экватор (рис. 4.1).
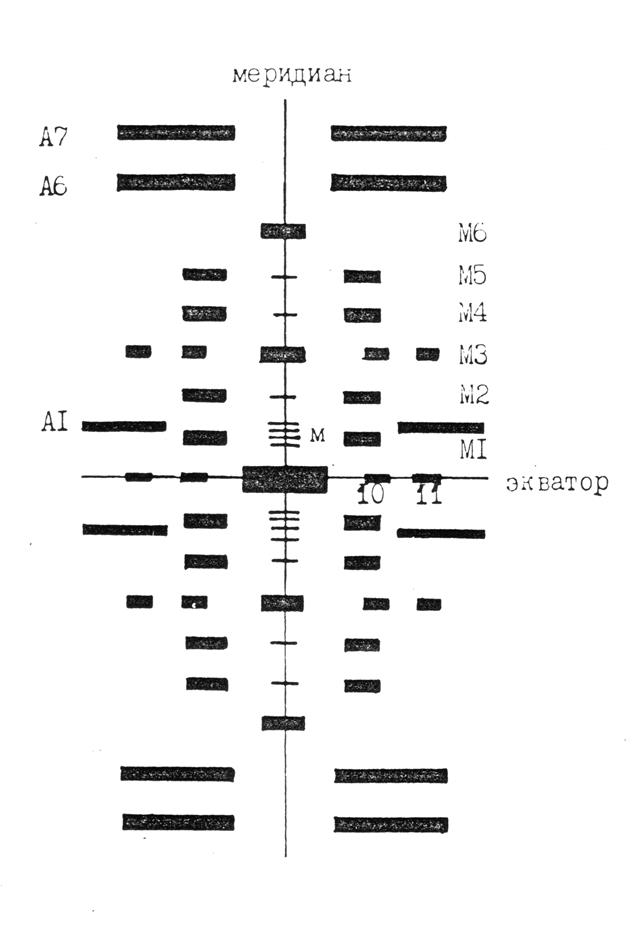
Схема рентгенограммы в достаточно грубом приближении представлена на рис. 4.2, где указаны два типа слоевых линий. Одна группа слоевых линий обусловлена дифракцией на актиновых (А), а другая – на миозиновых нитях (М). На миозиновых слоевых линиях интенсивность распределена и на меридиане, и вне его, а на актиновых – только вне меридиана.
|
|
Рис. 4.1. Малоугловая рентгенограмма живой
портняжной мышцы лягушки в состоянии покоя.
Видны слоевые линии. Образец размещался так, что хорошее
разрешение получалось только в меридиональном направлении
Кроме рефлексов, относящихся к слоевым линиям и обусловленных спиральным расположением компонентов толстых и тонких нитей, есть также «запрещенные» рефлексы, которые не должны бы были появиться, если бы дифракция происходила на спиральных структурах.
Экваториальное направление рентгенограммы имеет набор рефлексов (на рис. 4.2 обозначены индексами 10 и 11 два из них), характеризующих боковую упаковку толстых и тонких нитей в гексагональную решетку в области А-диска саркомера (рис. 4.3).
|
|
Рис. 4.2. Схема малоугловой рентгенограммы скелетной мышцы лягушки. М1, …, М6 – миозиновые слоевые линии (порядки периода 429 Å); А1, А6, А7 – актиновые слоевые линии (порядки периода 3´370 Å). Показаны некоторые так называемые «запрещенные» рефлексы, обозначенные м
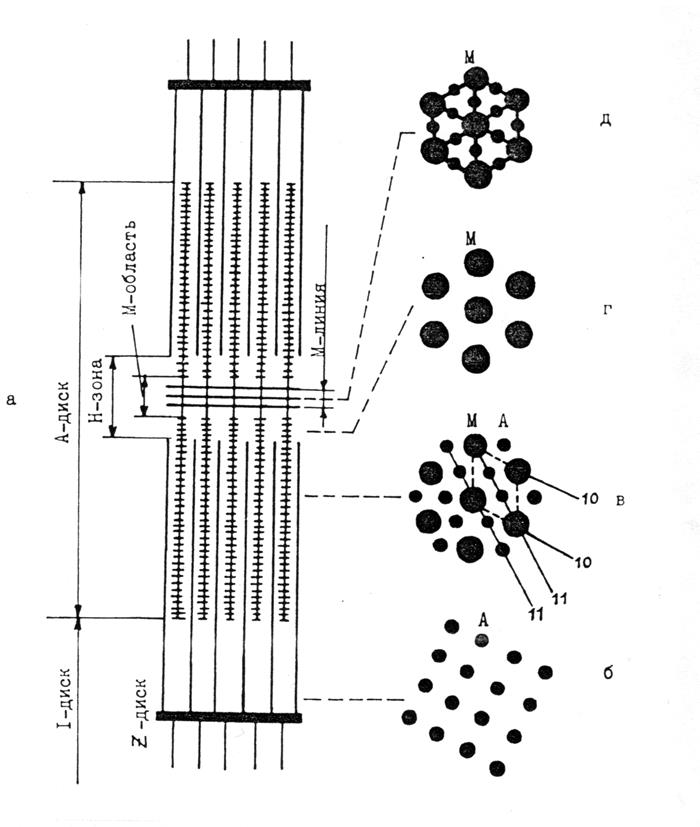
В этой решетке выделяют элементарную ячейку (рис. 4.3, в), которая содержит в углах толстые миозиновые нити (М), а в тригональных точках – тонкие актиновые нити (А).
|
|
Рис. 4.3. Схема саркомера скелетной мышцы, показывающая продольный вид (а) и виды в плоскостях, перпендикулярных к оси волокна: в I-диске вблизи Z-диска (б); в А-диске в области перекрытия толстых и тонких нитей (в); в Н-зоне (г) и в М-линии (д). В случае (в) показаны элементарная ячейка (пунктир) и плоскости (10) и (11) гексагональной решетки толстых и тонких нитей (М – миозиновые нити, А – актиновые нити)
Два наиболее интенсивных экваториальных рефлекса 10 и 11 соответствуют самым большим межплоскостным расстояниям и обусловлены соответственно отражением от плоскостей (10), включающих только толстые нити, и плоскостей (11), проходящих как через толстые, так и через тонкие нити (рис. 4.3, в). Интенсивности рефлексов 10 и 11 можно выразить через структурные факторы толстых и тонких нитей
![]() (4.1)
(4.1)
где ![]() и
и ![]() –
структурные факторы миозиновых и актиновых нитей. Соотношения (4.1) позволяют
оценить вклад толстых и тонких нитей в каждый экваториальный рефлекс. Можно
видеть, что изменения в величине
–
структурные факторы миозиновых и актиновых нитей. Соотношения (4.1) позволяют
оценить вклад толстых и тонких нитей в каждый экваториальный рефлекс. Можно
видеть, что изменения в величине ![]() больше влияют на
интенсивность рефлекса 10, чем на интенсивность рефлекса 11, тогда как к
увеличению или уменьшению величины
больше влияют на
интенсивность рефлекса 10, чем на интенсивность рефлекса 11, тогда как к
увеличению или уменьшению величины ![]() очень
чувствительна интенсивность рефлекса 11. Установлено, что рефлексы 10 и 11
обусловлены дифракцией на
очень
чувствительна интенсивность рефлекса 11. Установлено, что рефлексы 10 и 11
обусловлены дифракцией на ![]() -диске, а тонкие
нити упорядочены только в нем, причем упаковка тонких нитей меняется от тетрагональной
(рядом с
-диске, а тонкие
нити упорядочены только в нем, причем упаковка тонких нитей меняется от тетрагональной
(рядом с ![]() -диском) к гексагональной (в
-диском) к гексагональной (в ![]() -диске), а на большей части
-диске), а на большей части ![]() -диска они не имеют регулярного
расположения. Также было установлено, что при увеличении длины саркомера
происходит увеличение отношения интенсивностей
-диска они не имеют регулярного
расположения. Также было установлено, что при увеличении длины саркомера
происходит увеличение отношения интенсивностей ![]() и
и
![]() , что хорошо согласуется с моделью
скользящих нитей, лежащей в основе механизма сокращения.
, что хорошо согласуется с моделью
скользящих нитей, лежащей в основе механизма сокращения.
Экваториальные рефлексы 10 и 11 позволяют, в принципе, построить карты электронной плотности в проекции на плоскость, перпендикулярную к оси мышцы. При невысоком разрешении (~200 Å), которое получается, если используются эти два рефлекса при построении карты электронной плотности с помощью обратного преобразования Фурье (синтеза Фурье), можно считать структуру элементарной ячейки центросимметричной, что упрощает выбор фаз. По картам электронной плотности можно оценить относительные массы актиновых и миозиновых нитей.
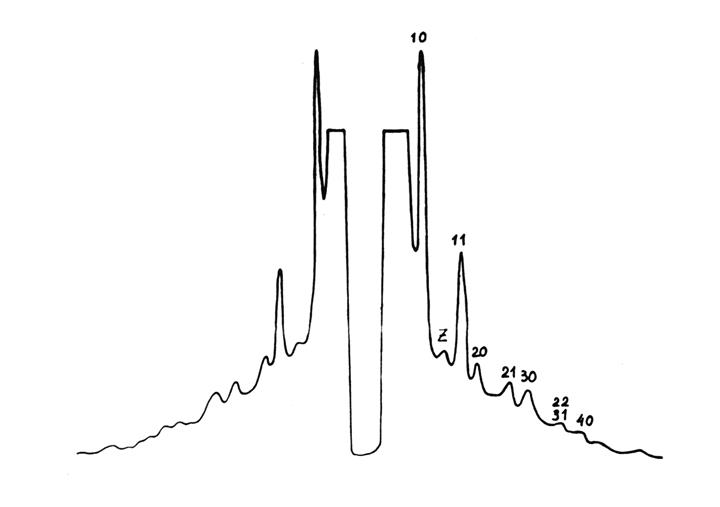
При использовании более усовершенствованной техники эксперимента на экваторе рентгенограммы живой мышцы наблюдаются рефлексы со значениями индексов h, k, большими, чем 10 и 11 (рис. 4.4). Учет этих рефлексов позволяет увеличить разрешение в картине электронной плотности, что способствует получению дополнительной информации. На этом пути есть известные трудности («проблема фаз»), которые, тем не менее, оказываются преодолимыми.
|
|
Рис. 4.4. Малоугловая рентгенограмма (денситограмма) живой
портняжной мышцы лягушки в состоянии покоя (экватор)
Кроме указанных выше рефлексов, которые хорошо объясняются дифракцией на гексагональной решетке толстых и тонких нитей, на экваторе рентгенограммы мышцы между рефлексами 10 и 11 регистрируется еще один рефлекс (на рис. 4.4 он обозначен буквой Z). По сравнению с рефлексами 10 и 11 он имеет существенно меньшую интенсивность и его межплоскостное расстояние слабо изменяется в зависимости от длины саркомера. Было показано, что основной вклад в указанный рефлекс дает структура Z-диска и примыкающих к нему тонких нитей, имеющих тетрагональную упаковку (рис. 4.3, б).
Рассмотрим далее интерпретацию слоевых линий рентгенограммы мышцы. Как уже отмечалось выше, имеются две системы слоевых линий. Одна из них, связанная с периодом 429 Å, обусловлена дифракцией на спиральной структуре, образованной поперечными мостиками толстых нитей.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.