Детали строения линии Z саркомера остаются дискуссионными в части прикрепления актиновых миофиламентов. Однако считают, что актиновые миофиламенты соседних саркомеров соединяются в области линии Z друг с другом посредством молекул, подобных белку тропомиозину, которые, начинаясь здесь, идут в направлении соседних саркомеров по актиновым миофиламентам. Кроме этого белка, в состав актиновых миофиламентов входит белок тропонин.
Актиновые миофиламенты имеют длину 1000 нм и диаметр от 4 до 8 нм, т. е. они короче и тоньше, нежели миозиновые. От линии Z актиновые миофиламенты проходят через половину диска I в диск А, располагаясь в последнем гексагонально вокруг миозиновых. В области гексагонального расположения миофиламентов, или в зоне перекрытия диска А, шесть актиновых миофиламентов окружают каждый миозиновый миофиламент, с которыми последний соединяется поперечными мостиками, образованными выступающими из толстого миофиламента частями молекул миозина. В расслабленном поперечнополосатом мышечном волокне актиновые миофиламенты противоположных сторон не доходят друг до друга в середине диска А на 0,2 мкм, и поэтому в центре последнего на фоне только одних миозиновых миофиламентов возникает светлая зона, или полоса Н (зона Хэнсона) (см. рис. 4).
Сокращению миофибрилл предшествует нервное возбуждение в области аксомышечного синапса, которое распространяется по Т-системе и достигает каждого саркомера. С Т-системы возбуждение передается на мембраны поперечной системы саркоплазматической сети, что ведет к увеличению проницаемости мембран для ионов кальция. Кальций регулирует взаимодействие мышечных белков в процессе мышечного сокращения. Энергию для мышечного сокращения доставляет гидролиз АТФ. Боковые расстояния между актиновыми и миозиновыми миофиламентами таковы, что они могут свободно скользить относительно друг друга в продольном направлении. Скольжение их обусловлено отклонением актиномиозиновых мостиков, при этом наблюдается смещение актиновых миофиламентов относительно миозиновых. Актиновые миофиламенты перемещаются в светлую зону саркомера на 0,1 мкм с каждой стороны. На первый взгляд укорочение саркомера на 0,2 мкм является малой величиной, однако если учесть, что миофибриллы состоят из большого количества саркомеров, то в целом достигается значительное сокращение поперечнополосатого мышечного волокна. Данная теория мышечного сокращения получила название теории скользящих нитей (рис. 6).
Поперечнополосатая скелетная мышечная ткань входит в состав мышц. Мышцу можно рассматривать как орган, поскольку ей присущи определенная структура и специфическая функция. Поперечнополосатые мышечные волокна объединяются при помощи соединительной ткани в орган, который и называется мышцей. Снаружи мышца покрыта соединительнотканной оболочкой — перимизием
.
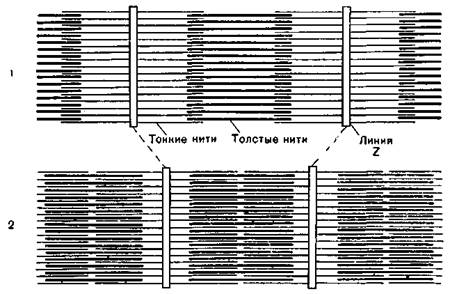
Рис. 6. Схема сокращения саркомера миофибриллы: / — строение саркомера в покое; 2 — то же при сокращении.
От последнего внутрь мышцы отходят прослойки рыхлой соединительной ткани, богатые кровеносными сосудами и нервами. Эти прослойки рыхлой соединительной ткани именуются эндомизием. Эндомизий располагается вокруг пучков поперечнополосатых мышечных волокон и отдельных мышечных волокон. Коллагеновые волокна эндомизия соединяются с сетью преколлагеновых волокон базальной мембраны поперечнополосатых мышечных волокон. На концах поперечнополосатых мышечных волокон сарколемма образует глубокие пальцеобразные выросты, в углублениях которых располагаются коллагеновые волокна сухожилий и фасций, с помощью которых мышца прикрепляется к скелету.
Поперечнополосатая сердечная мышечная ткань входит в состав стенок сердца. Основная масса миокарда состоит из поперечнополосатой сердечной рабочей мышечной ткани, которая отличается от поперечнополосатой скелетной мышечной ткани тем, что состоит не из поперечнополосатых мышечных волокон, а из мышечных клеток — миоцитов (см. рис. 7). Миоциты имеют прямоугольную форму, длина их достигает 50—120 мкм, а ширина — 15—20 мкм. В некоторых миоцитах наблюдается раздвоение конца клетки на две части, вследствие чего клетка приобретает У-образную форму. В центральной части миоцитов располагается одно, реже два ядра овальной или удлинен ной формы. Миофибриллы миоцитов в отличие от поперечнополосатых мышечных волокон располагаются не в центре клетки, а на периферии, они имеют толщину 1—3 мкм, и их строение подобно таковому поперечнополосатых мышечных волокон. Однако вследствие меньшего количества миофибрилл поперечная исчерченность миоцитов выражена слабее, чем поперечнополосатых скелетных мышечных волокон.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.