Серотонин присутствует в самых высоких концентрациях в тромбоцитах и в желудочно-кишечном тракте. Меньшие количества найдены в ядрах мозга (лимбическая система, новая кора) и сетчатке. После высвобождения из серотонинергических нейронов, большая часть высвобождаемого серотонина возвращается активно секретируемыми клетками. Некоторые антидепрессанты ингибируют этот механизм, способствуя более длительному пребыванию серотонина в синаптической щели.
Функция серотонина проявляется после его взаимодействия со специфичными рецепторами. Несколько рецепторов серотонина клонированы и идентифицированы как 5HT1, 5HT2, 5HT3, 5HT4, 5HT5, 5HT6, и 5HT7. В пределах этих групп выделяют подруппы( например группа 5HT1 имеет подгруппы 5HT1A, 5HT1B, 5HT1D, 5HT1E, и 5HT1F). Большинство этих рецепторов связаны с G белками, которые воздействуют на активность или аденилат циклазы или фосфолипазы Cg (PLCg). 5HT3 класс рецепторов представлен ионными каналами.
Некоторые рецепторы серотонина пресинаптические и другие постсинаптические. 5HT2A рецепторы опосредуют агрегацию тромбоцитов и сокращение гладких мышц. 5HT2C рецепторы связаны с механизмами всасывания в желудочно-кишечном транкте (показано, что мыши с дефектами гена этих рецепторов страдают ожирением из-за повышенного всасывания пищевых продуктов). 5HT3 рецепторы связаны с механизмами рвоты, а в регуляции секреции и перистальтики кишечника принимают участие 5HT4 рецепторы. 5HT6 и 5HT7 рецепторы широко распространены в лимбической системе мозга, а 5HT6 рецепторы имеют высокое сродство антидепрессантным препаратам
Мелатонин образуется из серотонина в эпифизе и сетчатке, в которых находится N-ацетилтрансфераза. Паренхиматозные клетки эпифиза секретирует мелатонин в кровь и цереброспинальную жидкость. Синтез и секреция мелатонина увеличиваются в течение темнового периода дня и поддерживаются в низком уровне в течение светлых часов. Эти суточные колебания синтеза мелатонина регулируются с участием норадреналина, секретируемого постганглионарными симпатическими нервами, иннервирующими эпифиз. Эффекты норадреналина проявляются через b - адренергические рецепторы, которые регулируют уровень цАМФ. Последняя через протеинкиназы активизирует N-ацетилтрансферазу, необходимую для синтеза мелатонина. Мелатонин в свою очередь ингибирует синтез и секрецию других медиаторов (дофамин и ГАМК).
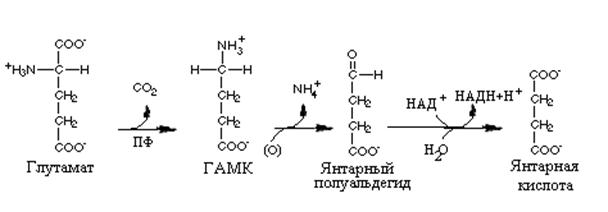 |
. Синтез и распад ГАМК
сером веществе головного мозга. Декарбоксилирование L-
глутамата - это основной путь биосинтеза g-аминомасляной кислоты. Возможно также ее образованием из путресцина (продукт дезаминирования орнитина).
Катаболизм g-аминобутирата начинается с потери аминогруппы и образования янтарного полуальдегида. Последний может быть восстановлен в g-гидроксибутират при участии L-лактатдегидрогеназы, либо окислиться с образованием янтарной кислоты и затем в цикле лимонной кислоты до СО2 и Н2О.
ГАМК оказывает тормозящий эффект на деятельность ЦНС. Ее препараты используют при лечении заболеваний , сопровождающихся возбуждением коры головного мозгпа
Гистамин играет важную роль во многих патологических процессах. Он образуется из гистидина путем декарбоксилирования. Эту реакцию катализирует декарбоксилаза ароматических L-аминокислот. Этот фермент не обладает выраженной субстратной специфичностью и катализирует также декарбоксилирование ДОФА, 5-гидрокситриптофана, фенилаланина, тирозина и триптофана. Декарбоксилаза in vitro и in vivo ингибируется а-метиламинокислотами, применяемыми в клинике в качестве гипотензивных средств. В большинстве клеток имеется также специфическая декарбоксилаза гистидина. Гистамин обладает широким спектром действия. Особенно хорошо известно его сосудорасширяющее действие. Он образуется локально в местах воспаления, обеспечивая доставку питательных веществ к месту повреждения, но может оказывать и общее действие, что часто приводит к резкому падению артериального давления.
Подобно другим биогенным аминам, гистамин разрушается путем окислительного дезаминирования при помощи моноаминоксидаз- флавинзависимых ферментов, локализованных преимущественно в митохондриях (МАО). Реакция необратима и протекает в два этапа. На первом этапе амин окисляется с передачей водородов на ФАД и образованием аммиака и соответствующего альдегида, а на втором этапе восстановленный кофермент окисляется молекулярным кислородом с образованием пероксида водорода. Ингибиторы МАО находят применение при лечении гипертонической болезни, депрессивных состояний и т.д.
Гистидин используется также в синтезе ряда небелковых соединений. К числу находящихся в организме гистидин содержащих соединений относятся эрготионеин (в эритроцитах и печени), карнозин и ансерин . 1-Метилгистидин, обнаруживаемый в моче человека, вероятно, образуется из ансерина. Содержание 3-метилгистидина в моче человека обычно составляет около 50 мг/100 мл, у пациентов с болезнью Вильсона оно значительно ниже.
Заключение.Таким образом в процессе катаболизма аминокислот происходят три типа реакций:.реакции по радикалу( сопровождающиеся процессами гидроксилирования, аминирования, сульфирования, фосфорилирования, изменения структуры радикала); второй тип реакций-декарбоксилирование с образованием биогенных аминов: третий тип- реакции дезаминирования ( прямого и непрямого), сопровождающиеся либо синтезом новых аминокислот, либо снятием аминогруппы с последующим включением углеродных скелетов в ЦТК или другие биосинтезы.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.