Гепатоциты расположенные вокруг капилляров портальной системы после приема пищи получают кровь, богатую питательными веществами. Эти клетки обогащены глутаминазой и ферментами синтеза мочевины. Глутаминаза аллостерически активируется ионами аммония, так что ионы аммония, поступающие в гепатоциты, активируют фермент, который превращает глутамин в ионы аммония и глутамат. Основная масса ионов аммония удаляется из циркуляции перипортальными гепатоцитами. Кm карбомоилфосфатсинтетазы для аммиака примерно 1 мM. Карбомоилфосфатсинтетаза I и глутаминаза аллостерически активируются N-ацетилглутаматом.
Перивенозные гепатоциты образуют один из слоев клеток, что окружают центральную вену. Эти клетки обогащены глутаминсинтетазой, с низким значением Кm для ионов аммония. Поэтому ионы аммония, которые не превратились в мочевину в перипортальных клетках, накапливаются в составе глутамина в перивенозных клетках.
Печень единственный
орган, клетки которого содержат все ферменты синтеза мочевины и, следовательно,
являются главным местом синтеза мочевины в организме. Ферменты, участвующие в
ее синтезе расположены в митохондриях и цитозоле . Синтез мочевины – пример циклического
метаболического пути. Орнитин представляет собой пример непротеиногенной
диаминомонокарбоновой кислоты, которая играет роль своеобразного катализатора
процесса, начиная этот процесс в качестве акцептора карбамоилфосфата и, в
заключительной реакции, после удаления молекулы мочевины она вновь
регенерирует. Расходуются в процессе мочевинообразования аспарагиновая кислота
и ион аммония ( источники аминогрупп молекулы мочевины), диоксид углерода (из
бикарбоната) и АТФ. Синтез 1 моля мочевины требует 3 молей АТФ.
. Синтез мочевины – пример циклического
метаболического пути. Орнитин представляет собой пример непротеиногенной
диаминомонокарбоновой кислоты, которая играет роль своеобразного катализатора
процесса, начиная этот процесс в качестве акцептора карбамоилфосфата и, в
заключительной реакции, после удаления молекулы мочевины она вновь
регенерирует. Расходуются в процессе мочевинообразования аспарагиновая кислота
и ион аммония ( источники аминогрупп молекулы мочевины), диоксид углерода (из
бикарбоната) и АТФ. Синтез 1 моля мочевины требует 3 молей АТФ.
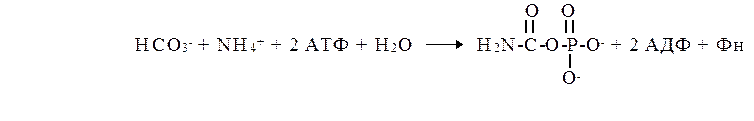 |
Карбамоилфосфат синтезируется в митохондриях в комплексной реакции, которая катализируется карбамоилфосфат синтетазой I. Этот аллостерический фермент активируется N –ацетилглутаминовой кислотой. Потребление 2 молекул АТФ делает синтез карбамоил-фосфата необратимой реакцией. Часть этой энергии депонируется в виде энергии макроэргической связи (смешанная ангидридная связь).
Карбамоилфосфат синтетаза I ( КФС-1) катализирует образование карбомоилфосата в митхондриях. Из цитозоля выделена карбомоилфосфатсинтетаза-2 (КФС-2), которая катализирует образование карбомоилфосфата, используемого для синтезе пиримидиновых нуклеотидов.
Цитруллин подобно орнитину - основная аминокислота, которая не кодируется генетически.
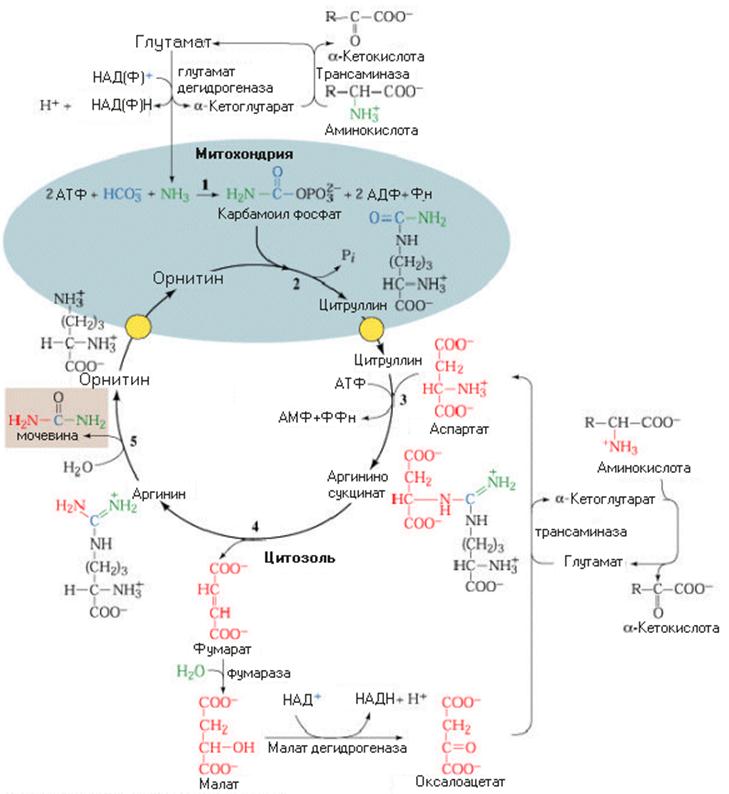
Орнитиновый цикл синтеза мочевины
Образование цитруллина происходит благодаря использованию высокоэнергетического фосфата карбамоилфосфата и катализируется орнитин-карбамоил трансферазой (ОКТ). Продукт реакции цитруллин транспортируется в цитозоль, где завершается образование мочевины.
Аминогруппа аспарагиновой кислоты - источник второго атома азота в молекуле мочевины. Конденсация аспарагиновой кислоты с цитруллином катализируется аргининосукцинат синтетазой (AС) и сопровождается гидролизом АТФ до АМФ и пирофосфата (ФФн). Так как пирофосфатазы быстро гидролизуют пирофосфат до 2 остатков фосфата, реакция сдвигается в направлении синтеза аргининосукцината.
Аргигин синтезируется при распаде аргининосукцината
Реакция распада аргининосукцината катализируется аргининосукцинат лиазой (АЛ) и образующийся аргинин является непосредственным предшественником мочевины. Фумаровая кислота, продукт этой реакции, позволяет связать этот процесс с другими путями, включая цикл Кребса, глюконеогенез и синтез заменимых аминокислот.
Распад аргинина обеспечивает образование мочевины и регенерирует орнитин
Гидролиз гуанидиновой группы аргинина катализируется ферментом аргиназой.Этот фермент в незначительных количествах обнаружен и в других органах (почки, мозг и др.), но наиболее активен в печени. Мочевина путем диффузии по градиенту концентрациии поступает в кровь и транспортируется к почкам, где фильтруется и выделяется. Около 25% ее поступает в кишечник, где она становится субстратом для уреазы. Другой продукт реакции орнитин переносится вновь митохондрии и участвует в следующем туре цикла синтеза мочевины.
Суммарная реакция синтеза мочевины
![]() Аспарагиновая кислота + NH3 + CO2 +
3АТФ
Аспарагиновая кислота + NH3 + CO2 +
3АТФ
![]() 3 H2O + мочевина + фумарат + 2АДФ +
АМФ + 2Фн + пирофосфат
3 H2O + мочевина + фумарат + 2АДФ +
АМФ + 2Фн + пирофосфат
Между циклом мочевинообразования и циклом трикарбоновых кислот устанавливается тесная связь на уровне субстратов (см рис.).
Один атом азота доставляется ионом аммония, а другой - аспарагиновой кислотой. Глутаминовая кислота - непосредственный предшественник и аммиака, и аспартата.
Синтез мочевины – регулируемый процесс
Быстрая регуляция синтеза мочевины происходит на уровне КФС-1.Этот фермент аллостерически регулируется N-ацетилглутаминовой кислотой. Последняя синтезируется в митохондриях из глутамата и ацетил-КоА. Реакция катализируется при помощи синтазы, которая в свою очередь активируется аргинином.
![]() Ацетил-КоА + глутамат N-ацетилглутаминовая
кислота + КоА
Ацетил-КоА + глутамат N-ацетилглутаминовая
кислота + КоА
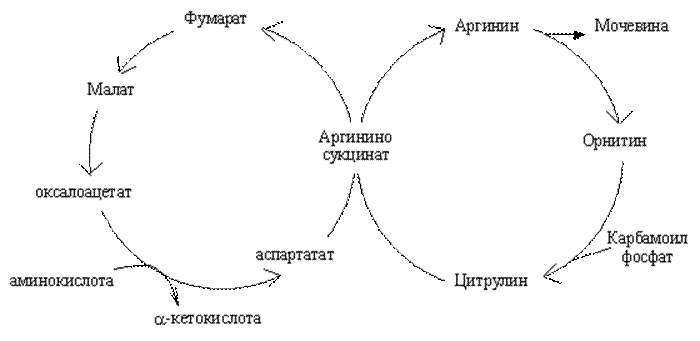
Связь орнитинового и лимоннокислого циклов на уровне субстратов.
Уровень N -ацетилглутаминовой кислоты зависит от соотношения активностей N-ацетилглутамат синтазы и N-ацетилглутамат гидролазы, которая катализирует гидролиз этого соединения.
Долговременная регуляция направлена на синтез новых ферментов. Индукция синтеза определяется уровнем пищевого белка. Повышение поступления белков с пищей повышает синтез всех ферментов синтеза мочевины. Это показано в следующей таблице:
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.