Подобная регуляция осуществляется с помощью ДНК-связывающих белков, один из которых активирует, а другой ингибирует транскрипцию.
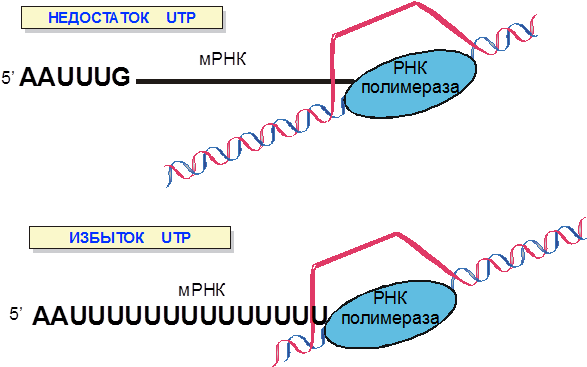
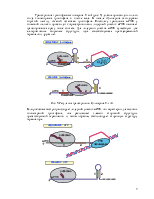
Рис. 7. Регуляция транскрипции пиримидинового оперона: реитеративная инициация транскрипции.
Кроме замены s-факторов и использования белков-регуляторов ряд оперонов регулируется третьим способом, реитеративной транскрипцией. Известно, что при инициации транскрипции первые несколько полимеризованных нуклеотидов плохо связаны в транскрипционном комплексе. Эта особенность используется при регуляции транскрипции пиримидинового оперона. Если концентрация UTP в клетке высока, нет нужды транскрибировать этот оперон. В начале оперона расположен короткий участок, кодирующий олигоуридиловую кислоту. При высокой концентрации UTP олигоU фрагмент начинает проскальзывать по ДНК, так, что синтезируется длинные олигоуридиловые фрагменты, что приводит к терминации транскрипции (рис. 7).
Кроме пиримидинового оперона, опероны рибосомных РНК также регулируются в зависимости от концентрации нуклеотидтрифосфатов. Промоторы генов рРНК оказываются очень чувствительными к уровню инициирующих нуклеотидов – ATP и GTP. Таким образом, при слишком высокоинтенсивной трансляции или в условиях недостатка энергии, когда существенно понижается уровень ATP и GTP, транскрипция генов рРНК приостанавливается.
Кроме стадии инициации, транскрипция может регулироваться также с помощью преждевременной терминации, т.е. в зависимости от нужд клетки должен осуществляться выбор между элонгацией и терминацией. Наиболее простой способ остановить транскрипцию – связать белок-регулятор на пути РНК-полимеразы. Так происходит в случае оперона, ответственного за синтез пуриновых нуклеотидов. Два более сложных механизма регулируемой терминации это антитерминация и аттенюация.
Аттенюация – это регулируемая терминация зависящая от относительной стабилизации альтернативных вторичных структур транскрибируемой мРНК. Как известно, r-независимый терминатор представляет собой РНК шпильку и последующий олигоуридиловый участок. Если будет стабилизирована альтернативная вторичная структура, где олигоуридиловый фрагмент удален от ближайшей шпильки, терминация не будет происходить. Есть несколько факторов, стабилизирующих одну из альтернативных вторичных структур РНК во время транскрипции.
Транскрипция триптофанового оперона E. coli (рис. 8) должна происходить только когда концентрация триптофана в клетке мала. В начале trp-оперона закодирован короткий пептид, богатый остатками триптофана. Поскольку трансляция мРНК у кишечной палочки происходит котранскрипционно, лидерный участок мРНК начинает транслироваться сразу после синтеза. Для лидерного участка мРНК существуют две альтернативные вторичные структуры, одна способствующая преждевременной терминации, другая нет.
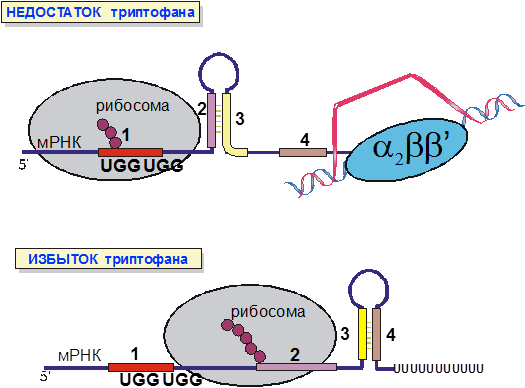
Рис. 8. Регуляция транскрипции trp-оперона E. coli.
Если рибосома быстро транслирует лидерный участок мРНК, что характерно для высоких концентраций триптофана, она расплавляет элемент вторичной структуры, препятствующий терминации и, таким образом, стабилизирует вторичную структуру терминатора.
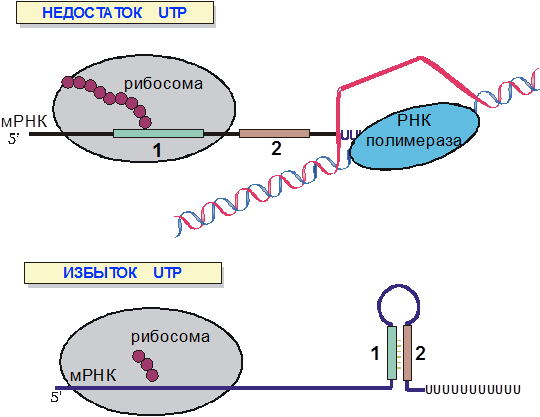
Рис. 9. Регуляция транскрипции пиримидинового оперона с помощью аттенюации.
Если в регуляции транскрипции trp-оперона кишечной палочки выбор между альтернативными вторичными структурами мРНК определяется скоростью трансляции, то для пиримидинового оперона, решающий фактор скорость транскрипции (рис. 9). Если уровень UTP велик, то РНК-полимераза быстро прочитывает олигоуридиловую последовательность регулируемого терминатора и транслирующая мРНК рибосома не успевает приблизиться к шпильке терминатора. Если концентрация UTP низка, т.е. в условиях, когда экспрессировать оперон необходимо, РНК-полимераза замедляется при синтезе олигоуридилата. При этом транслирующая рибосома догоняет РНК-полимеразу и препятствует образованию шпильки терминатора.
Наконец, самый простой способ стабилизировать одну из альтернативных вторичных структур РНК используется для регуляции транскрипции trp-оперона B. subtilis (рис.10). В присутствии больших концентраций триптофана триптофан-связывающий белок TRAP взаимодействует с транскрибирующейся мРНК trp-оперона и стабилизирует конформацию, способствующую преждевременной терминации.
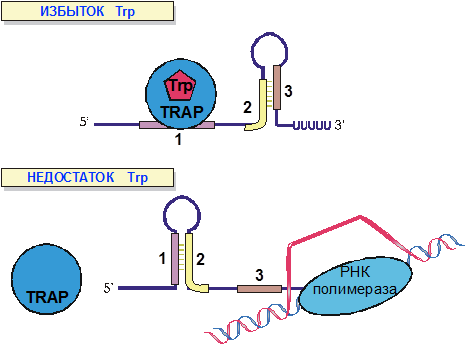
Рис. 10. Регуляция транскрипции триптофанового оперона B. subtilis с помощью аттенюации.
Кроме аттенюации, регуляция терминации также происходит при т.н. антитерминации. Лучше всего процесс антитерминации изучен для бактериофага
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.