В мембране микроворсинок
кишечника расположены специфические транспортные системы, участвующие в
активном транспорте аминокислот в энтероциты. Различают несколько таких систем
для переноса: 1. нейтральных аминокислот, 2. - алифатических гидрофобных
аминокислот, 3. - иминокислот, 4. - кислых аминокислот, 5. - основных
аминокислот. Работа некоторых их этих переносчиков зависит от ионов натрия,
градиент которого (как и при всасывании глюкозы) обеспечивает их перенос через
мембрану. Два переносчика также требуют Cl-. У двух
систем транспорт независим от Na+.
Ди- и
трипептиды транспортируются в энтероциты системой, которая требует Н+
вместо Na+. Большие
пептиды всасываются в очень небольших количествах. В энтероцитах олигопептиды
распадаются до аминокислот внутриклеточным гидролизом и образовавшиеся
аминокислоты вместе с аминокислотами, посту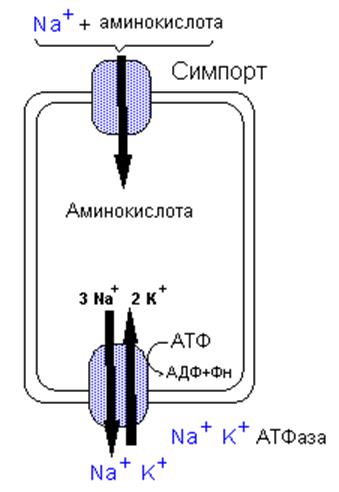 пившим из
кишечника покидают клетку через базолатеральную мембрану при помощи по крайней
мере пяти транспортных систем. Две из этих систем зависимы и три независимы от
Na+.
Из межклеточного пространства аминокислоты и небольшие пептиды попадают в
систему воротной вены.
пившим из
кишечника покидают клетку через базолатеральную мембрану при помощи по крайней
мере пяти транспортных систем. Две из этих систем зависимы и три независимы от
Na+.
Из межклеточного пространства аминокислоты и небольшие пептиды попадают в
систему воротной вены.
Всасывание аминокислот активно происходит в верхних отделах тонкого кишечника. Часть белка не усваивается в тонком кишечнике и поступает в толстый кишечник, где используется микрофлорой.
У новорожденных возможно всасывание умеренных количеств непереваренных белков. Антитела материнского молока - представленные секреторными иммуноглобулинами (IgA), поступают в кровь из кишечника при помощи эндоцитоза с последующим экзоцитозом и обеспечивают пассивный иммунитет против инфекций. Этот процесс снижается с возрастом , но взрослые все еще абсорбируют небольшие количества белка. Чужеродные белки поступающие в кровь, способствуют образованию антител, и реакция на последующее поступление большего количества того же белка может вызвать аллергические симптомы, что объясняет происхождение аллергических реакций после приема некоторых пищевых продуктов.
Поглощение белковых антигенов, например, бактериальных и вирусных белков, про
Поглощение белковых антигенов, например, бактериальных и вирусных белков, происходит в больших М клетках, специализированных кишечных эпителиоцитах, которые передают антигены к лимфоидным клеткам, и активируют лимфобласты, в последующем секретирующие IgA в ответ на поступление того же антигена. Этот секреторный иммунитет - важный механизм защиты в кишечнике.
Определенная, иногда значительная часть белков и аминокислот попадает в толстый кишечник и используется микроорганизмами кишечника в качестве источника аминокислот для синтеза белков и энергетических субстратов. Превращения аминокислот под влиянием микрофлоры кишечника получили название гниения белков. Многие продукты, образуемые бактериями для человека являются токсическими и могут представлять клинический интерес. Основные химические процессы, лежащие в основе гниения связаны с реакциями декарбокислирования, дезаминирования, десульфирования и удаления боковой части ароматических аминокислот.
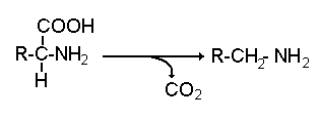 В
результате действия бактериальных декарбоксилаз образуется целый ряд биологически
активных аминов. Так, при декарбоксилировании лизина образуется диамин
кадаверин (трупный яд), при декарбоксилировании тирозина- тирамин, орнитина –
путресцин, гистамин образуется из гистидина. Многие из этих продуктов
вазоактивные вещества.
В
результате действия бактериальных декарбоксилаз образуется целый ряд биологически
активных аминов. Так, при декарбоксилировании лизина образуется диамин
кадаверин (трупный яд), при декарбоксилировании тирозина- тирамин, орнитина –
путресцин, гистамин образуется из гистидина. Многие из этих продуктов
вазоактивные вещества.
 |
Толстый кишечник является источником значительных количеств аммиака, образующегося при дезаминировании аминокислот. Обычно этот аммиак после всасывания обезвреживается печенью, но при заболеваниях печени уровень аммиака в периферической крови может достигать токсических значений. Кровотечения в кишечник или прием больших количеств белка у таких больных может приводить к аммиачной интоксикации.
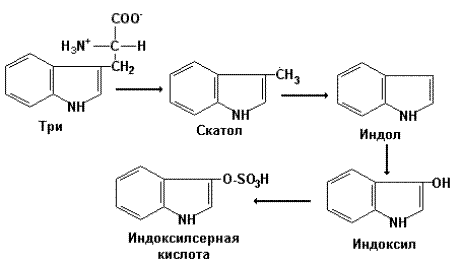
Особое место в образовании токсических продуктов занимает обмен ароматических аминокислот. Под влиянием декарбоксилаз, дезаминаз и других ферментов бактерий происходит постепенное укорочение боковых цепей ароматических аминокислот и остающиеся циклические части этих аминокислот, обладая выраженными гидрофобными свойствами могут оказывать токсическое влияние на клетки печени после их всасывания.
Так, при постепенном удалении боковой цепи триптофана образуются скатол и индол, тирозина –крезол и фенол. После всасывания эти продукты взаимодействуют с активными формами серной (арилсульфотрансфераза) или глюкуроновой (УДФ-глюкуронилтрансфераза) кислот с образованием парных кислот, растворимых в воде и выделяемых почками. Количество этих продуктов в моче может отражать выраженность процессов гниения в кишечнике.
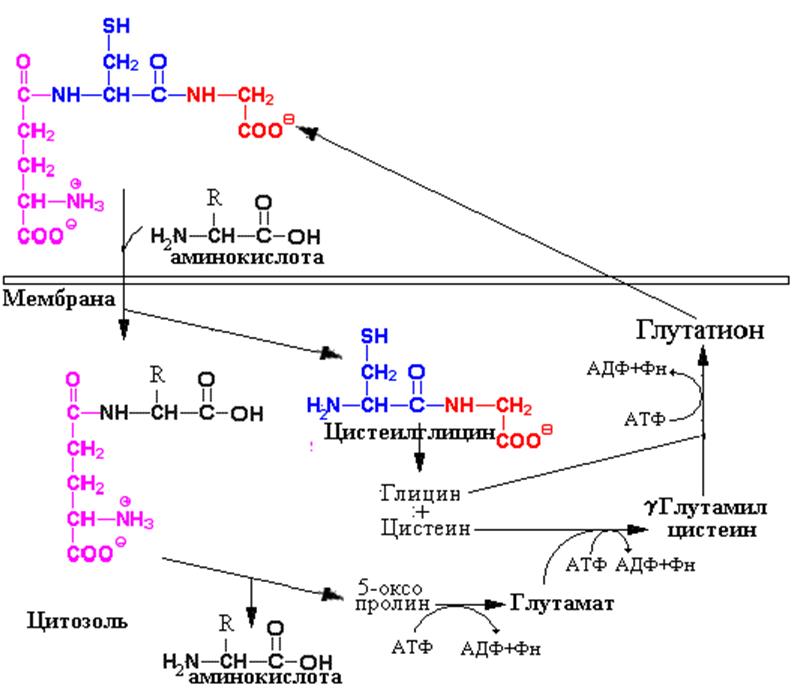
Одним из механизмов переноса аминокислот через клеточные мембраны является процесс, получивший название g-глутамиловый цикл. В основе его лежит реакция катализируемая g- -глутамилтранспептидазой Этот ферментявляется структурным компонентом мембран и обеспечивает взаимодействие глутатиона с поступающей в клетку аминокислотой. Реакция с аминокислотой высвобождает цистеил-глицин из глутатиона, а образующийся дипептид (g-глутамил-аминокислота) транспортируется в клетку и гидролизуется там с высвобождением переносимой аминокислоты. Глутамат при этом превращается 5-оксопролин, а цистеил-глицин распадается до аминокислот. Последующие реакции включают энергозависимую регенерацию глутатиона, включающую превращение 5-оксопролина в глутаминовую кислоту (1 моль АТФ)и синтез трипептида в две последовательные реакции (2 моля АТФ). Этот процесс, таким образом, требует больших энергетических затрат, но обладает высокой скоростью и емкостью. Наиболее активен этот механизм при реабсрорбции аминокислот эпителием канальцев почек.
 |
Организационно- Методические указания:
1.Мультимедийная презентация
доцент каф. Свергун В.Т.
Дата
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.