Жирные кислоты с очень длинной цепью. Особенностью метаболизма жирных кислот в пероксисомах является расщепление тех из них, которые имеют очень длинную углеводородную цепь или другие необычные радикалы, неспособные подвергаться эффективному окислению в митохо ндриях. Укорочение алкильной цепи в пероксисомах происходит до тех пор, пока не образуется ацил-КоА со средней длиной цепи. Это обусловлено субстратной специфичностью пероксисомальной ацил-КоА дегидрогеназы. Образующийся ацил-КоА с С-8 впоследствии подвергается дальнейшему окислению в митохондриях.
Первоначальная стадия дегидрирования в ходе пероксисомального окисления жирных кислот протекает с образованием Н2О2, а не ФАДН2. Перекись водорода удаляется с помощью каталазы. Все последующие реакции аналогичны происходящим в митохондриях, хотя катализируются они изоферментами пероксисом.
Окисление дикарбоновых кислот. В пероксисомах происходит также окисление дикарбоновых кислот, образующихся в ходе w-окисления. Само w-окисление протекает в эндоплазматическом ретикулуме и занимает малую долю в окислительных процессах, которым подвергаются жирные кислоты. При w-окислении гидроксилирование происходит на метильном конце жирнокислотной цепи; в результате образуется дикарбоновая кислота.
Окисление жирных кислот с разветвленной цепью. Растения и молочные продукты содержат фитановую кислоту. Это жирная кислота, имеющая в своем составе 20С и 4 метильные группы. Окисление таких жирных кислот становится возможным в организме благодаря пероксисомальному ферменту - a-гидроксилазе. Недостаток этого фермента сопровождается развитием болезни Рефзума.
Болезнь Рефзума. При болезни Рефзума в тканях и жидкостях организма накапливается
фитановая кислота - необычная жирная кислота с разветвленной цепью. Она образуется из фитола, который поступает в организм с растительной пищей, богатой хлорофиллом. В норме фитановая кислота в пероксисомах подвергается a-окислению. В ходе этого процесса она укорачивается на один углеродный атом с образованием пристановой кислоты и СО2. Пристановая кислота, подвергаясь b-окислению, расщепляется на 3 молекулы ацетил-КоА, 3 молекулы пропионил-КоА и 1 молекулу изобутирил-КоА. У людей с болезнью Рефзума имеется генетический дефект образования рецептора, обусловливающего пероксисомальную локализацию фермента - фитанат гидроксилазы. Интересно, что последующая стадия превращения пристановой кислоты при этом не нарушена. Клиническая симптоматика у таких больных связана с развитие пигментной дегенерации сетчатки, хронической полинейропатии и мозжечковой атаксии, повышением концентрации белка в цереброспинальной жидкости.
Незаменимые и заменимые - Среди ненасыщенных жирных кислот в организме человека не могут синтезироваться w-3 и w-6 жирные кислоты в связи с отсутствием ферментной системы, которая могла бы катализировать образование двойной связи в положении w-6 или любом другом положении, близко расположенном к w-концу. К таким жирным кислотам относятся линолевая кислота (18:2, D9,12), линоленовая кислота (18:3, D9,12,15) и арахидоновая кислота (20:4, D5,8,11,14). Последняя является незаменимой только при недостатке линолевой кислоты, поскольку в норме она может синтезироваться из линолевой кислоты.
У человека при недостатке в пище незаменимых жирных кислот описаны дерматологические изменения. Обычный рацион взрослых людей содержит достаточное количество незаменимых жирных кислот. Однако у новорожденных, которые получают рацион, обедненный жирами, отмечаются признаки поражения кожи. Они проходят, если в курс лечения включается линолевая кислота. Случаи подобного дефицита наблюдаются и у пациентов, которые длительное время находятся на парентеральном питании, обедненном незаменимыми жирными кислотами. В качестве профилактики такого состояния достаточно, чтобы в организм поступали незаменимые жирные кислоты в количестве 1-2% от общей калорической потребности.
Синтез ненасыщенных жирных кислот из насыщенных с параллельным удлинением цепи. Десатурация проходит под действием микросомального комплекса ферментов, состоящего из трех компонентов белковой природы: цитохрома b5, цитохром b5-редуктазы и десатуразы, которые содержат в своем составе негемовое железо. В качестве субстратов используются НАДФН и молекулярный кислород. Из этих компонентов образуется короткая цепь переноса электронов, с помощью которой на короткий период времени в молекулу жирной кислоты включаются гидроксильные группы. Затем они отщепляются в виде воды, в результате в молекуле жирной кислоты формируется двойная связь. Имеется целое семейство субъединиц десатуразы, которые специфичны к определенному месту введения двойной связи.
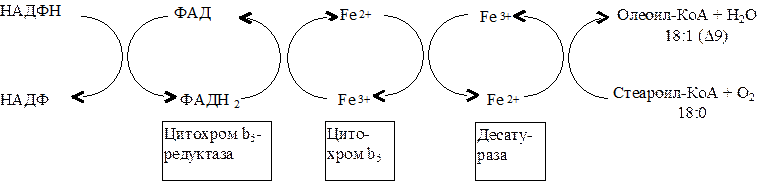
Микросомальный комплекс ферментов и система транспорта электронов
Элонгация происходит путем добавления двухуглеродного фрагмента к карбоксильному концу жирной кислоты. Имеется две раздельных системы элонгации в микросомах и митохондриях. В микросомальной системе элонгации в качестве донора двухуглеродной группировки используется малонил-КоА, а в митохондриальной системе - ацетил-КоА. Введение двухуглеродного фрагмента включает две реакции восстановления (используются 2 молекулы НАДФН) и последовательность других реакций, описанных в разделе “синтез жирных кислот”.
Двумя основными видами ацетоновых тел являются ацетоацетат и b-гидроксибутират. b-гидроксибутират - это восстановленная форма ацетоацетата. Ацетоацетат образуется в клетках печени из ацетил~КоА. Образование происходит в митохондриальном матриксе.
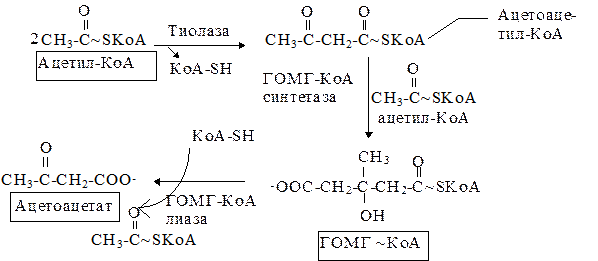
Первоначальная стадия этого процесса катализируется ферментом - b-кетотиолазой. Затем ацетоацетил-КоА конденсируется со следующей молекулой ацетил-КоА под влиянием фермента ГОМГ-КоА синтетазы. В результате образуется b-гидрокси-b-метилглютарил-КоА. Затем фермент - ГОМГ-КоА лиаза катализирует расщепление ГОМГ-КоА на ацетоацетат и ацетил-КоА. В дальнейшем ацетоуксусная кислота восстанавливается под влиянием фермента b-гидроксибутиратдегидрогеназы и в результате образуется b-оксимасляная кислота. Количество ацетоацетата, которое восстанавливается в b-гидроксибутират, зависит от соотношения НАДН/НАД+. Восстановление это происходит под влиянием фермента b-гидроксибутиратдегидрогеназы. Печень служит главным местом образования кетоновых тел благодаря высокому содержанию ГОМГ-КоА синтетазы в митохондриях гепатоцитов.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.