В растворах могут существовать различные механизмы возникновения разности потенциалов: диффузионные, мебранные и межфазные.
В первом случае возникновение разности потенциалов связано с различием в скоростях диффузии положительных и отрицательных ионов.
Во втором – с существованием между растворами селективно проницаемой мембраны. В этом случае механизм возникновения контактной электродвижущей силы подобен механизму, имеющему место в металлах и полупроводниках: одна группа ионов вследствие градиентов концентраций может переходить границу раздела растворов, другая – нет. Происходит разделение зарядов, степень которого определяется возникающей разностью потенциалов.
И, наконец, третий механизм связан с отличием в подвижности ионов в различных фазах одного и того же вещества. При контакте фаз ионы в неодинаковой степени осуществляют межфазный переход, что приводит к возникновению областей с отличающимися концентрациями зарядов и, следовательно, разности потенциалов.
В процессе развития исследований для объяснения природы биопотенциалов использовались все три механизма электрогенеза.
В настоящее время достаточно точно установлено, что основной механизм возникновения потенциала между цитоплазмой и межклеточной жидкостью связан с селективной проницаемостью клеточной мембраны. Селективная проницаемость обусловлена существованием в мембране пор – каналов, сформированных определенными белками, пронизывающими мембрану. Тип белка, формирующего канал, определяет и его избирательную проницаемость. Существуют натриевые, калиевые, кальциевые и другие каналы. Селективность каналов, однако, не абсолютна. Например, проницаемость натриевого канала аксона кальмара для Na+, Li+, K+, Rb+, Cs+ соотносится как 1:1,1:0,083:0,025:0,016, а калиевого для той же группы элементов – как 0,010:0,018:1:0,910:0,077. Основными ионами, определяющими мембранный потенциал в различных клетках, являются ионы калия, натрия, кальция и хлора, вследствие чего в отмеченных пропорциях важны лишь отношения между этими ионами. Как видно, проницаемость натриевого канала для натрия в 12 раз превышает проницаемость для калия, а проницаемость калиевого канала для калия в 100 раз превосходит проницаемость для натрия. Проницаемости каналов нервных и мышечных клеток, однако, не остаются постоянными. Под действием возбуждающего фактора они могут сильно изменяться и становиться функцией времени и мембранного потенциала. Число ионных каналов на 1 мкм2 в мембране варьирует в широких пределах – от десятков в нервных волокнах позвоночных до 500 в аксоне кальмара. Размеры каналов составляют порядок единиц ангстрем. К примеру, натриевые каналы имеют прямоугольное сечение с размерами сторон 0,31х 0,51 нм, а калиевые – квадратные – 0,45х0,45 нм. Размеры ионов при этом составляют 0,266 нм для иона калия и 0,196 нм для иона натрия, т.е. имеют, в общем-то, тот же порядок, что и размеры каналов, вследствие чего в канале практически происходит их однорядное движение. Для получения представления об интенсивности движения ионов в каналах можно привести данные по пропускной способности одиночного натриевого канала. Она имеет порядок 107 ионов/с. Мембранные каналы работают независимо друг от друга.
Установлено, что для каналов
различных ионов существуют вещества, блокирующие их работу. При этом блокатор
каналов одного типа ионов может не влиять на проницаемость каналов другого типа
ионов. Ионные каналы возбудимых мембран обладают свойством изменения
проводимости. При этом проводимость отдельного канала изменяется скачкообразно
от непроводящего в проводящее состояние. Канал открывается с определенной
вероятностью на некоторое время. Эта вероятность является функцией мембранного
потенциала. Каналы каждого типа ионов характеризуются временем жизни, т.е.
временем, в течение которого вероятность их открывания велика. Для натриевых
каналов это время составляет единицы миллисекунд. Вследствие скачкообразного
изменения тока отдельного канала суммарный ток, протекающий через мембрану,
должен был бы иметь ступенчатый вид. Однако при большом числе каналов этого не
происходит. Поясним. Определим относительную флюктуацию тока мембраны. Ток I мембраны, обусловленный действием N
каналов, определяется суммой токов отдельных каналов ![]() :
:
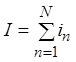 . Среднее значение тока мембраны
. Среднее значение тока мембраны ![]() равно сумме средних токов
равно сумме средних токов ![]() отдельных каналов
отдельных каналов 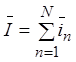 . Если все каналы одинаковы, то
одинаковы и их средние токи
. Если все каналы одинаковы, то
одинаковы и их средние токи ![]() . Тогда
. Тогда ![]() .
.
Относительная флюктуация мембранного тока K равна
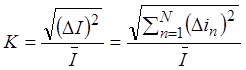 ,
,
где DI – отклонения от среднего мембранного тока, ![]() - отклонения от среднего тока
канала.
- отклонения от среднего тока
канала.
Так как каналы
одинаковы, средние значения квадратов отклонений токов каналов тоже одинаковы,
т.е. 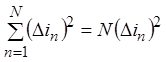 и
и 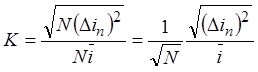 .
.
Поскольку канал
переходит из открытого в полностью закрытое состояние, то можно приближенно
считать ![]() . Тогда
. Тогда ![]() .
.
Из этого соотношения следует, что по мере увеличения числа каналов, участвующих в токопереносе, относительная флюктуация уменьшается. Обычно суммарное число каналов в мембране велико, и поэтому, несмотря на скачкообразный характер токов отдельных каналов, ток мембраны изменяется плавно.
Резюмируя, можно отметить следующие основные свойства мембранных каналов:
1) селективность;
2) независимость работы каналов;
3) вероятностный характер проводимости.
Кроме этого, проводимости отдельных каналов возбудимых мембран проявляют нелинейные и параметрические свойства.
Перемещения ионов по каналам под действием градиентов концентраций (пассивный транспорт) приводят к выравниваю плотностей зарядов по обе стороны мембраны. Для поддержания разности концентраций между внутренней средой клетки и межклеточной жидкостью существуют механизмы переноса ионов из области с меньшей концентрацией в область с большей (активный транспорт). Одним из основных способов переноса зарядов против градиентов концентраций является транспортировка с помощью так называемых ионных насосов. Роль насосов исполняют молекулы специальных белков, встроенных в мембрану. В настоящее время известны три типа насосов: калий-натриевый (K+-Na+), кальциевый (Ca++) и протонный (Н+). Работа против градиентов требует затрат энергии. В качестве ее источника служит химическое соединение, именуемое аденозинтрифосфорной кислотой (АТФ). Работа ионных насосов осуществляется за счет уменьшения свободной энергии этой молекулы.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.