При малых t изменение U незначительно. Вследствие этого уровень активации натриевых каналов оказывается малым и ток ионов натрия не достигает уровня калиевого тока. К моменту же, когда потенциал мембраны возрастает до существенных значений, начинается процесс инактивации натриевых каналов, в результате чего происходит уменьшение натриевого тока. Таким образом, в этой ситуации условие (18) не выполняется и возбуждение не происходит.
Рефрактерность. Если сразу после генерации потенциала действия на мембрану подать деполяризующий импульс, то в течение определенного времени она остается нечувствительной к нему, даже если уровень тока значительно превышает пороговый. В дальнейшем чувствительность мембраны постепенно восстанавливается и пороговый уровень от больших значений в начале появления регенеративной реакции уменьшается до стационарного значения. Интервал времени, в течение которого чувствительность мембраны понижена или вообще отсутствует, называется рефрактерным периодом. Время полного отсутствия чувствительности называется абсолютным рефрактерным периодом, а интервал ослабленной чувствительности – относительным рефрактерным периодом.
Явление рефрактерности объясняется
следующим образом. Спад возбуждения мембраны связан с инактивацией натриевых
каналов. Вследствие этого их интегральная проводимость оказывается весьма малой
и нечувствительной к изменениям напряжения на мембране. Активность калиевых
каналов в это время еще сохраняется и весьма значительна. Поэтому при
деполяризации мембраны натриевый ток практически не изменяется, а калиевый
возрастает, и, следовательно, даже если выполняется условие (18), условие (19)
не выполняется (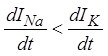 ). По мере уменьшения
степени инактивации натриевый ток начинает увеличиваться, и при достаточном
стимуле условия (18) и (19) могут оказаться выполненными, т.е. произойдет
очередной регенеративный процесс.
). По мере уменьшения
степени инактивации натриевый ток начинает увеличиваться, и при достаточном
стимуле условия (18) и (19) могут оказаться выполненными, т.е. произойдет
очередной регенеративный процесс.
1.3.3. Электрическая модель возбудимой мембраны
Процессы, протекающие в возбудимых мембранах при формировании потенциала действия, подобны процессам в релаксационных генераторах, в частности релаксаторах радиоэлектронной природы. Релаксационные генераторы характеризуются устойчивыми и квазиустойчивыми состояниями. Устойчивым называется состояние, в котором система может находиться сколь угодно долго в отсутствие внешних сил, квазиустойчивым – состояние, в котором система пребывает лишь в течение конечного времени, определяемого внутренними параметрами. В зависимости от соотношения состояний релаксационные генераторы делятся на три группы: автоколебательные, с внешним запуском и триггеры. Первые характеризуются отсутствием устойчивых состояний и совершают постоянные переходы между двумя квазиустойчивыми. Вторые (называемые также ждущими) имеют одно устойчивое и одно квазиустойчивое состояния, причем переход из первого во второе осуществляется под действием внешней силы, а возвращение – вследствие внутренних свойств. Третьи могут находиться в одном из двух устойчивых состояний, переходы между которыми также происходят при внешнем воздействии.
Сравнивая поведение биологической мембраны возбудимой клетки с приведенной классификацией, можно видеть, что активная мембрана относится к типу релаксационных систем с внешним запуском – способна сколь угодно долго находиться в состоянии покоя и генерировать потенциал действия при появлении внешнего стимула достаточной амплитуды. Длительность формируемого импульса зависит от типа биологической структуры: потенциал действия аксона кальмара имеет длительность около 1 мс, клетки скелетной мышцы – 2-3 мс, клетки сердечной мышцы (кардиомиоцита) – 250-300 мс.
Одна из многочисленных возможных моделей возбудимой мембраны представлена на рис. 19. Устройство представляет собой мультивибратор с внешним запуском. Собственно мультивибратор построен на транзисторах Т3 и Т4. Каскады на транзисторах Т1 и Т2 позволяют наиболее полно моделировать процесс возбуждения и сохранять полярность стимулирующего импульса такой же, как у биологического аналога. Каскад на транзисторе Т2 имитирует каналы ионных токов утечки. Токи возбужденного состояния воспроизводятся каскадом на транзисторе Т3. Конденсатор См – эквивалентная емкость мембраны.
В отсутствие внешнего импульса
транзистор Т4 открыт и насыщен током базы, протекающим через
резистор R7, а транзистор Т3
закрыт. Закрытое состояние Т3 обеспечивается тем, что запирающее напряжение
между его базой и эмиттером, создаваемое падением напря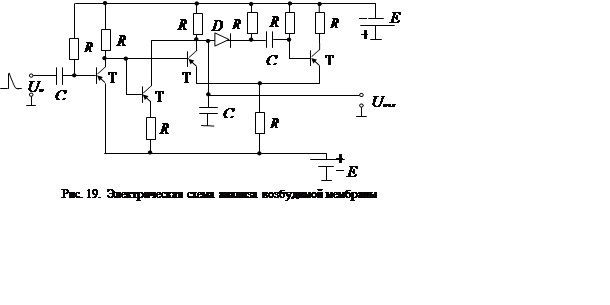
жения на резисторе R6 вследствие
протекания через него тока эмиттетра транзистора Т4, превышает
отпирающее напряжение, равное разности потенциалов между коллектором и
эмиттером транзистора Т1, являющегося одним из плеч делителя в цепи
базы Т3. Это условие можно записать в виде: ![]() ,
где
,
где ![]() - ток эмиттера транзистора Т4,
- ток эмиттера транзистора Т4,
![]() - падение напряжения на транзисторе
Т1. Транзистор Т2 находится в активном режиме. В его
коллекторной цепи протекает ток
- падение напряжения на транзисторе
Т1. Транзистор Т2 находится в активном режиме. В его
коллекторной цепи протекает ток ![]() . Вследствие этого
потенциал коллектора Т3 равен алгебраической сумме
. Вследствие этого
потенциал коллектора Т3 равен алгебраической сумме ![]() и имеет отрицательный знак (E2 < 0). Конденсатор
и имеет отрицательный знак (E2 < 0). Конденсатор ![]() заряжен до напряжения
заряжен до напряжения ![]() . Заряд его происходит через
резисторы R5 и R6
и переход эмиттер-база транзистора Т4. Диод D
исключает из цепи заряда
. Заряд его происходит через
резисторы R5 и R6
и переход эмиттер-база транзистора Т4. Диод D
исключает из цепи заряда ![]() резистор R4, чтобы не происходило влияние зарядного
тока на характер изменения мембранного потенциала. Источник Е2
имитирует равновесный калиевый потенциал, Е1 – равновесный
натриевый.
резистор R4, чтобы не происходило влияние зарядного
тока на характер изменения мембранного потенциала. Источник Е2
имитирует равновесный калиевый потенциал, Е1 – равновесный
натриевый.
В этом состоянии,
которое может сохраняться сколь угодно долго, напряжение на выходе устройства (![]() ) равно
) равно ![]() .
Это напряжение имитирует потенциал покоя. После подачи на вход устройства
импульсного напряжения так же, как и в случае биологической мембраны, возможны
различные варианты в зависимости от амплитуды, полярности и формы импульса.
Рассмотрим некоторые из них.
.
Это напряжение имитирует потенциал покоя. После подачи на вход устройства
импульсного напряжения так же, как и в случае биологической мембраны, возможны
различные варианты в зависимости от амплитуды, полярности и формы импульса.
Рассмотрим некоторые из них.
1. Запускающий импульс прямоугольной формы имеет деполя-ризующую (положительную) полярность. Амплитуда импульса ниже пороговой (ниже уровня возникновения регенеративного процесса).
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.