На меридиане малоугловой рентгенограммы мышцы регистрируются рефлексы, которые, как отмечено выше, не объясняются дифракцией на спиральных структурах толстых и тонких нитей (рис. 4.7). Некоторые из этих рефлексов связывают со структурой толстых нитей, а другие относят
|
|
Рис. 4.6. Схема образования дифракционной картины от спирали тонкой нити. Эту спираль, имеющую, например, 13 субъединиц в 6 оборотах (в), можно представить как результат умножения непрерывной спирали с периодом 59 Å (а) и набора плоскостей, отстоящих друг от друга на 27,5 Å (б). [В (а) и (в) показаны радиальные проекции спиралей]. Дифракционная картина такой спирали (е) образуется в результате свертки дифракционных картин от непрерывной спирали (г) и от набора плоскостей (д)
к дифракции на тонких нитях. При интерпретации меридиональных
рефлексов основываются на том факте, что миозиновые и актиновые нити упакованы
в структуры, состоящие из двух половин. Например, М-линия делит А-диск
на две части, а для тонких нитей такой границей служит Z-диск,
если рассматривать участок, состоящий из двух ![]() -дисков
соседних саркомеров с Z-диском между ними (рис.
4.3). Это обстоятельство приводит к тому, что происходит интерференция
излучений, рассеянных каждой половиной. Такую структуру можно представить как
свертку какой-либо структурной картины одной половины с решеткой, состоящей из
двух точек, разделенных некоторым расстоянием (периодом интерференционной
функции). Тогда по теореме свертки искомая дифракционная картина будет произведением
дифракционных картин этих составляющих свертки (две точки и какая-либо структура
А- или
-дисков
соседних саркомеров с Z-диском между ними (рис.
4.3). Это обстоятельство приводит к тому, что происходит интерференция
излучений, рассеянных каждой половиной. Такую структуру можно представить как
свертку какой-либо структурной картины одной половины с решеткой, состоящей из
двух точек, разделенных некоторым расстоянием (периодом интерференционной
функции). Тогда по теореме свертки искомая дифракционная картина будет произведением
дифракционных картин этих составляющих свертки (две точки и какая-либо структура
А- или ![]() -дисков). Таким образом,
удалось объяснить многие так называемые «запрещенные» рефлексы на меридиане малоугловой
рентгенограммы мышцы.
-дисков). Таким образом,
удалось объяснить многие так называемые «запрещенные» рефлексы на меридиане малоугловой
рентгенограммы мышцы.
|
|
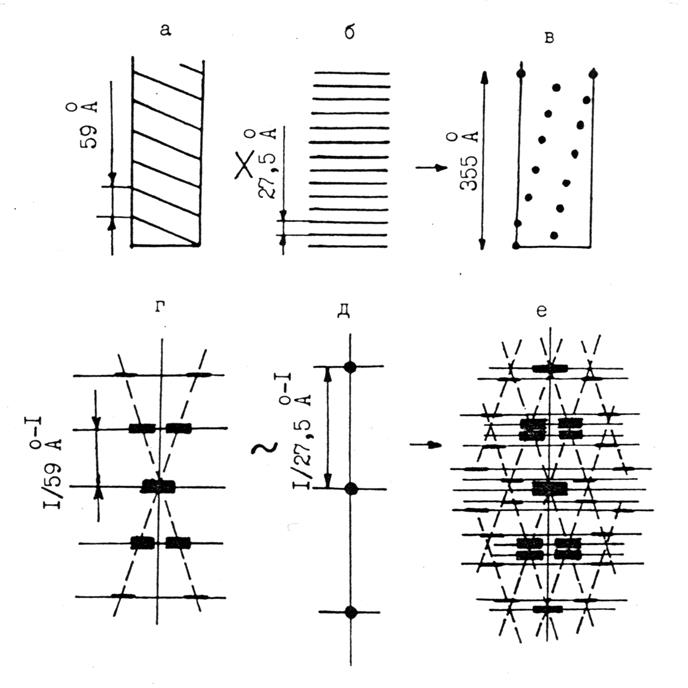
Рис.4.7. Малоугловая рентгенограмма живой покоящейся
скелетной мышцы (меридиан), на которой видны меридиональные рефлексы, не соответствующие дифракции на спиральных
структурах актиновых и миозиновых нитей
Выше были описаны некоторые характерные особенности рентгенограмм живых мышц в состоянии покоя. Однако, кроме таких мышц, рентгенографически изучены сокращающиеся мышцы и мышцы в состоянии ригора (трупного окоченения). Как следует из этих данных, и в том и в другом случае происходят значительные изменения в малоугловых рентгенограммах.
Рассмотрим вначале переход мышцы в ригор. В этом случае на экваторе
рентгенограммы происходит перераспределение интенсивностей рефлексов 10 и 11.
Если для живой покоящейся мышцы ![]() , то для
аналогичного образца, переведенного в ригор,
, то для
аналогичного образца, переведенного в ригор, ![]() ,
где
,
где ![]() и
и ![]() –
интенсивности рефлексов 10 и 11 (рис. 4.8).
–
интенсивности рефлексов 10 и 11 (рис. 4.8).
|
|
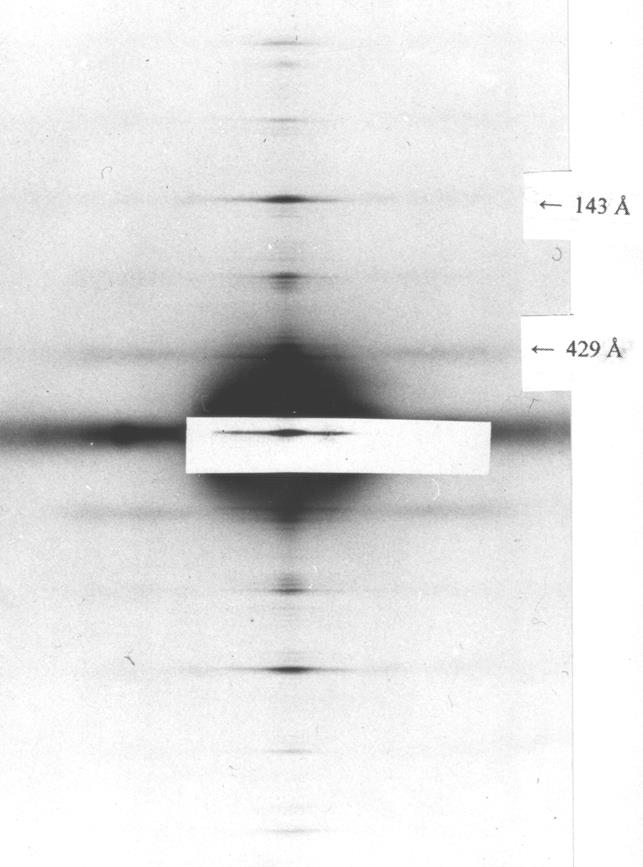
Рис. 4.8. Малоугловые рентгенограммы скелетной мышцы (экватор)
в состояниях покоя (а) и ригора (трупного окоченения) (б)
Такой результат был объяснен тем, что при переходе в ригор происходит перенос массы поперечных мостиков от толстых нитей к тонким, так что в ригоре мостики присоединены к актиновым нитям. При переходе в ригор также существенно перераспределяется интенсивность слоевых линий. Например, полностью исчезают миозиновые слоевые линии, характерные для рентгенограммы живой покоящейся мышцы, и появляются новые слоевые линии двух типов, межплоскостные расстояния которых соответствуют порядкам отражений периода 725 Å (рис. 4.9).
|
|
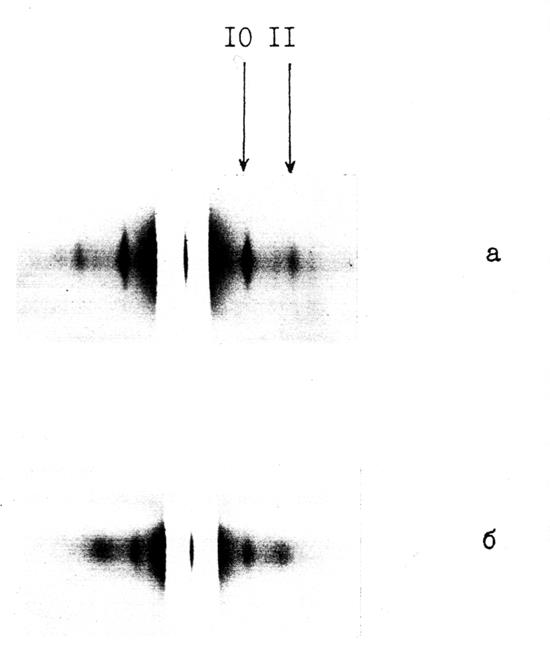
Рис. 4.9. Малоугловая рентгеногамма скелетной мышцы
(слоевые и меридиан) в состоянии ригора. Наблюдается значительное изменение картины слоевых линий по сравнению
со случаем живой покоящейся мышцы (см. рис. 4.1)
Кроме этого увеличивается интенсивность актиновых рефлексов, а также происходят некоторые другие изменения. Не вдаваясь в детали, укажем, что все эти изменения слоевых линий могут быть также объяснены присоединением поперечных мостиков к тонким нитям, так что мостики «усиливают» их структуру и располагаются в соответствии с симметрией тонкой нити, имеющей период спирали 360-370 Å. Причем эти данные хорошо согласуются с изменениями экваториальных рефлексов при переходе мышцы в ригор, указанными выше.
Изучение сокращающихся мышц осложняется тем, что время одиночного изометрического сокращения достаточно мало (~1 с). Соответственно, чтобы получить несколько рентгенограмм в процессе сокращения, необходимо регистрировать малоугловые рентгенограммы за очень малые промежутки времени (миллисекунды). Поэтому на ранних стадиях рентгенографического изучения под малыми углами сокращающихся мышц получение рентгенограмм осуществлялось путем многократного (с определенным интервалом) повторения коротких тетанических сокращений (часто от нескольких образцов) и суммирования экспозиций в каждом сокращении (в период максимального развития напряжения) с целью накопления необходимого экспозиционного времени. Позже в технику эксперимента были введены позиционно-чувствительные детекторы рентгеновского излучения, а вместо рентгеновских трубок стало использоваться синхротронное излучение. Данные компоненты в технике малоугловых рентгенографических исследований мышц существенно (на 2-4 порядка) сократили время, необходимое для получения дифракционной картины мышцы. Все это позволило рентгенографически изучать мышечное сокращение с высоким временным разрешением (миллисекунды).
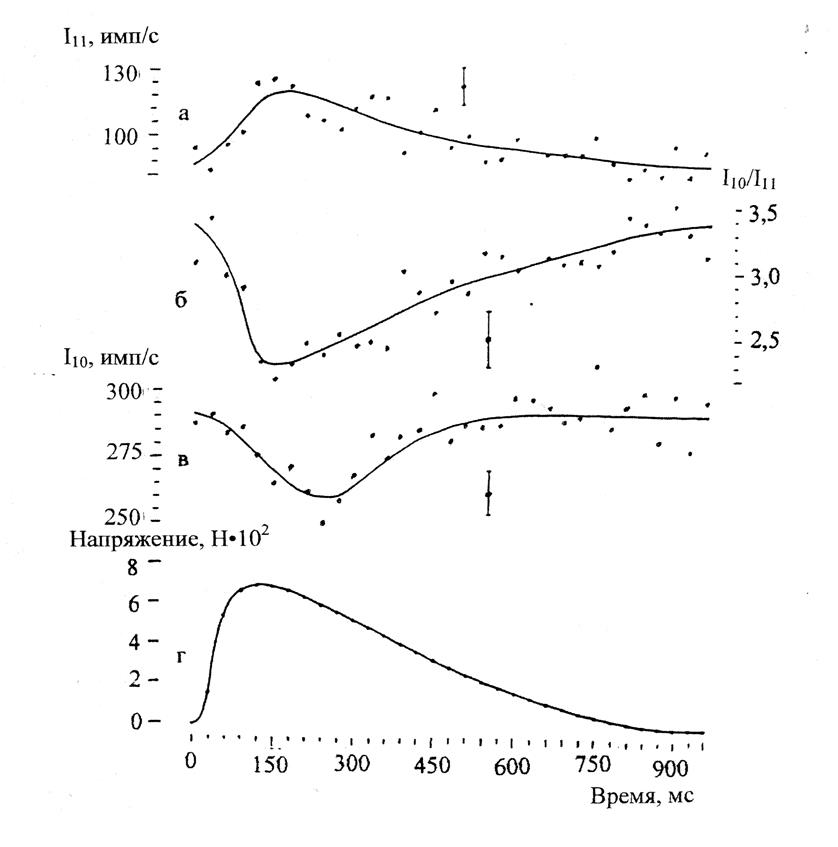
Изучались те же группы рефлексов, о которых уже шла речь выше. Было установлено, например, что ход изменений интенсивностей рефлексов 10 и 11 хорошо коррелирует с изменением напряжения, развиваемого изометрически сокращающейся мышцей (рис. 4.10), что подтверждало мостиковую схему механизма генерации силы при сокращении. Из остальных рефлексов остановимся только на актиновых слоевых линиях. Было обнаружено, что интенсивность актиновой слоевой линии 59 Å увеличивается при сокращении, но это увеличение меньше, чем в случае ригора. Это также свидетельствовало в пользу механизма мышечного сокращения по типу скользящих нитей с участием мостиков между ними.
|
Рис. 4.10. Зависимости от времени интенсивностей рефлексов 10 (в) и 11 (а), их отношения I10/I11 (б) и напряжения, развиваемого мышцей, во время одиночного изометрического сокращения. Временное разрешение 30 мс. Данные получены при использовании синхротронного излучения, малоугловой фокусирующей камеры и однокоординатного детектора |
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.