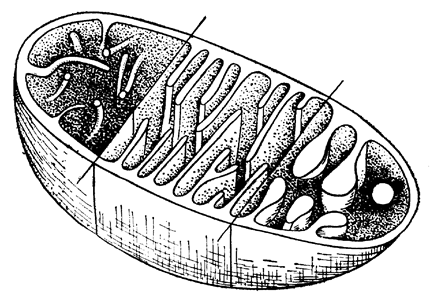
В 1978 г. Тэйлор (Taylor) сообщил, что в пределах каждой "хорошей" группы простейших строение крист митохондрий не меняется. Возникла гипотеза, что эукариот следует делить на две основные ветви - носителей митохондрий с трубчатыми кристами и носителей митохондрий с пластинчатыми кристами (рис. 2, 3). До сих пор совершенно непонятно, почему строение крист митохондрий обладает таким исключительным консерватизмом; кроме того, оказалось, что форма крист митохондрий эукариот более разнообразна, чем это виделось Тэйлору. В результате в западноевропейской и североамериканской научной литературе система Тэйлора не получила полного развития и здесь существует тенденция сохранения царства Простейшие. Однако отечественные ученые, в первую очередь Серавин (1980), Старобогатов (1986) и Мирабдуллаев (1989) и. Более осторожно, Кусакин и Дроздов (1994, 1997) продолжили это направление. Предлагаемая автором система ближе к системе Тэйлора в редакции Мирабдуллаева.
Концепция "молекулярных часов" позволяет выявлять родственные связи и строить филогенетические деревья без обращения к палеонтологии.
|
Рис. 3. Полусхематическое изображение трех различных типовкрист митохондрий. Слева: трубчатый, в центре - пластинчатый, справа - переходный, который может быть следствием искажений при подготовке препарата. Из Либберта, 1976 |
В 80-х годах появилась возможность исследовать различия в последовательностях кодонов гомологичных генов различных групп простейших. Теоретический аппарат концепции "молекулярных часов" - нейтралистская ("недарвиновская") теория эволюции Кимура и Ота* к этому времени была основательно разработана, а биохимические методы достаточно совершенны, чтобы реализовать ее возможности.
Основные данные для проверки макросистем эукариот получены на последовательностях кодонов рибосомной РНК (поскольку рибосомы имеются во всех типах живой клетки, начиная от древнейших прокариот). Банки данных о структуре рибосомной РНК постоянно растут. Рибосомы удивительно консервативны, поэтому количество (а в некоторых случаях и качество) различий в последовательностях их кодонов напрямую свидетельствуют о том, как давно разошлись сравниваемые группировки. Кладограммы (схемы, показывающие точки ветвления филогенетического дерева), построенные на анализе структуры рибосом, позволяют решить многие спорные вопросы систематики, однако методы построения кладограмм достаточно сложны, поэтому результаты, полученные с использованием разных методик или разных участков рРНК, зачастую сильно различаются. Однако бесспорно, что рРНК хроматофоров прокариотного типа и близка к рРНК сине-зеленых водорослей, а рРНК митохондрий свидетельствует об их происхождении от пурпурных бактерий. Эволюционные схемы, полученные с помощью этого метода, не опровергают целостности таксонов, выделенных по строению крист, хотя допускают и иные варианты системы.
Новая группа: Архезоа- простейшие, не имеющие митохондрий и пластид
Простейшие, не имеющие митохондрий, были известны сравнительно давно. Отсутствие митохондрий может быть объяснено либо тем, что это древнейшие реликты "домитохондриальной" эпохи развития эукариот, сохранившиеся в тех биотопах, где нет кислорода, либо вторичным исчезновением митохондрий как приспособлением к местообитаниям, где нет или мало кислорода. Основные группы безмитохондриальных простейших (микроспоридии и парабазалии) являются паразитами, а у паразитов, даже из числа многоклеточных, часто происходит выпадение некоторых ферментов окислительного цикла - они могут использовать промежуточные продукты обмена хозяина. На основании анализа последовательностей рРНК было окончательно доказано, что в основном своем большинстве эти животные (в том числе микроспоридии и парабазалии) являются исключительно древними. Их рибосомы структурно ближе к рибосомам прокариот, чем к рибосомам прочих эукариот и они же обладают наиболее примитивными формами митоза. Ныне безмитохондриальные эукариоты с 70S-рибосомами объединяются в царство Archezoa.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.