III постулат: центральный (главный) постулат гласит, что передача информации идет только в направлении: ДНК è РНК è белок и обратная передача информации невозможна. Этот постулат является центральной догмой всей биологии: как классической, так и современной молекулярной биологии. Для классической биологии – это стена, разделяющая дарвинизм и классическую генетику (учение Менделя-Моргана) от ламаркизма и лысенковщины, так как только изменения генотипа, ДНК (мутации), могут вызвать изменения фенотипа (белков тела, ферментов и т.д.), но не наоборот. Согласно Ламарку – благоприобретенные изменения (тела, признаков, т. е. белков) передаются по наследству (изменяют гены, ДНК).
Сюда относят следующие компоненты, необходимые и достаточные для обеспечения процесса биосинтеза белка (рис. 4.1) не только в клетке (как про-, так и эукариотической), но и вне клетки, в искусственно созданных условиях (и in vitro):
1. м-РНК;
2. рибосома;
3. аминоацил-т-РНК;
4. ГТФ;
5. белковые трансферные факторы:
5.1. факторы инициации;
5.2. факторы элонгации;
5.3. факторы терминации.
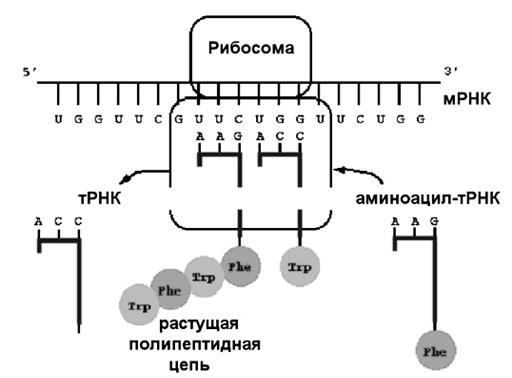
Рис. 4.1 Схема биосинтеза белка.
Не следует забывать еще и о факторах, создающих строго определенные физико-химические параметры среды (рН, ионная сила раствора, определенная концентрация катионов магния и т.д.).
Как и всякая РНК, она синтезируется в ходе транскрипции на матрице ДНК из дезоксирибонуклеозидтрифосфатов с помощью фермента ДНК-зависимой РНК-полимеразы по принципу комплементарности к одной из цепочек ДНК, называемой смысловой (значащей). Синтез м-РНК включает этапы: инициация, элонгация и терминация.
Инициация начинается с определенного участка – промотора. Его местоположение определяется специальными регуляторными молекулами. После того, как промотор узнается индукторами, он освобождается от репрессии, и обеспечивается его транскрипция. При этом синтезируются гигантские РНК (пре-м-РНК). После синтеза наступает этап созревания (процессинг), который заключается в разрезании синтезированной молекулы РНК на фрагменты, деградация акцепторной (регуляторной) зоны, интронов. Затем наступает этап сплайсинга – склеивания необходимых экзонов и формирование необходимой для трансляции последовательности нуклеотидов – м-РНК, созревание которой завершается кэпированием с 5'-конца («надевается шапочка») и полиаденилированием с 3'-конца (присоединение полиаденилового «хвоста»). Часто при содействии специальных белков образуется информосома и м-РНК транспортируется через поры ядерной оболочки в цитоплазму для трансляции.
Как уже упоминалось, рибосомы – это цитоплазматические немембранные органеллы, обеспечивающие синтез белков и состоящие из двух субъединиц, синтезированных в ядрышке и транспортированных в цитоплазму. Нормальное исходное для биосинтеза белка состояние рибосом – это свободные субъединицы, взвешенные в цитозоле (гиалоплазме) у эукариот или в миксоплазме у прокариот, способные связываться с белковыми факторами инициации и взаимодействовать своим М-центром (маленькие субъединицы) с м-РНК.
Процесс объединения т-РНК с аминокислотами называется активацией аминокислот. Он происходит в гиалоплазме благодаря наличию специальных ферментов аминоацил – т-РНК-синтетаз в два этапа:
1. Взаимодействие аминокислоты с ферментом (за счет пространственной специфичности радикала аминокислоты к структуре фермента), сопровождаемое гидролизом молекулы АТФ. Ее энергия используется для активации комплекса. В результате этой реакции образуется промежуточное соединение – аминоациладенилат.
2. За счет специфичности другого центра фермента к антикодону т-РНК происходит замена АМФ на т-РНК, но только ту, антикодон которой соответствует специфичности фермента. Таким образом, образуется специфический комплекс аа-т-РНК (аминоацил-т-РНК) ферментом аминоацил-т-РНК-синтетаза.
Уважаемый посетитель!
Чтобы распечатать файл, скачайте его (в формате Word).
Ссылка на скачивание - внизу страницы.